A. Degree of expansion
A defining moment in embryonic development is when fluid starts to accumulate between cells at the morulae stage of development. As the fluid's volume increases, a cavity appears gradually forming the blastocoel. This normally happens between Days 4 and 5 in human embryos in vitro and marks a new ‘era’ in the embryo's life, the blastocyst stage. As the fluid inside the newly formed blastocyst increases, so does the number of cells, and the combination of these two features causes a progressive enlargement of the blastocyst and it's cavity with a consequent progressive thinning of the zona pellucida (ZP). Finally, the blastocyst breaks free of the ZP through a process called hatching. The number of cells that comprise a blastocyst can vary considerably as shown in one study to range between 24 and 322 cells (Hardarson et al., 2003), which is often reflected in the blastocyst's morphology. The physiological events that underlie this transformation of a ‘cellular mass’ at the compaction stage to a highly structured blastocyst are not fully understood. However, cells that either by chance or fate are located in the outer part of the embryo start to flatten out, making contact with neighboring cells through tight junctions. In this way a barrier is created between the outside and the inside of the embryo, a prerequisite for blastocyst formation. Blastocyst formation is initiated through an initial secretion between the morula cells and this small cavity is then maintained and increased by actions of the membrane channels Na/K-ATPase that raise the salt concentration within the embryo, attracting water through osmosis (Watson et al., 2004). This increased water pressure gradually increases the size of the cavity which continues throughout the blastocyst stages.
The Istanbul consensus document (Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology, 2011) uses a simplified system of only four groups combining the first two and the last two groups of the Gardner and Schoolcraft (1999) grading system into two single groups which may be a limitation for the possibility to assess blastocysts. In this Atlas we have adopted the grading system that divides the grade of expansion into six categories and the ICM (Section B) and TE cell (Section C) grading into three categories similar to Gardner and Schoolcraft (1999) but have used the numerical scoring system suggested by the Istanbul consensus document: Grade 1 blastocysts are those in which the blastocoel cavity is less than half of the volume of the embryo (Figs 303–308); Grade 2 blastocysts are those in which the blastocoel cavity is half, or more than half, of the volume of the embryo (Figs 309–314); Grade 3 blastocysts are those in which the blastocoel cavity completely fills the embryo (Figs 315–320); Grade 4 blastocysts in which the blastocyst cavity is now greater than the original volume of the embryo and the ZP is thinned (Figs 321–326); Grade 5 blastocysts or hatching blastocysts in which the blastocoel cavity is greater than the original volume of the embryo and the TE is herniating through a natural breach in the ZP (Figs 327–332) and Grade 6 blastocyts or hatched blastocysts are those in which the blastocyst has completely escaped from a natural breach in the ZP (Figs 333–338). The latter two grades should be distinguished from blastocysts that are hatching or have hatched from an artificial breach in the ZP created by an assisted hatching procedure or following embryo biopsy whereby the breach in the ZP is quite large, permitting the blastocyst to escape earlier and well before complete expansion of the blastocoel cavity. The artificially hatched blastocyst could therefore contain far fewer cells than those that undergo natural hatching.
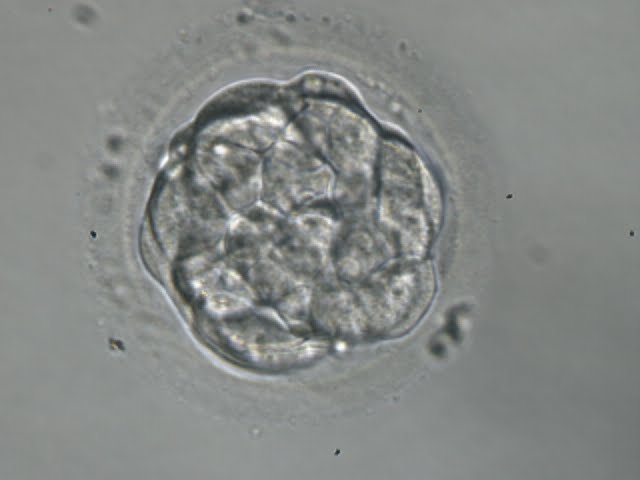
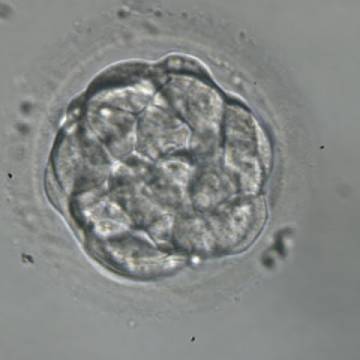
Figure 303
A very early blastocyst with a small cavity appearing centrally. The blastocyst was transferred but the outcome is unknown.
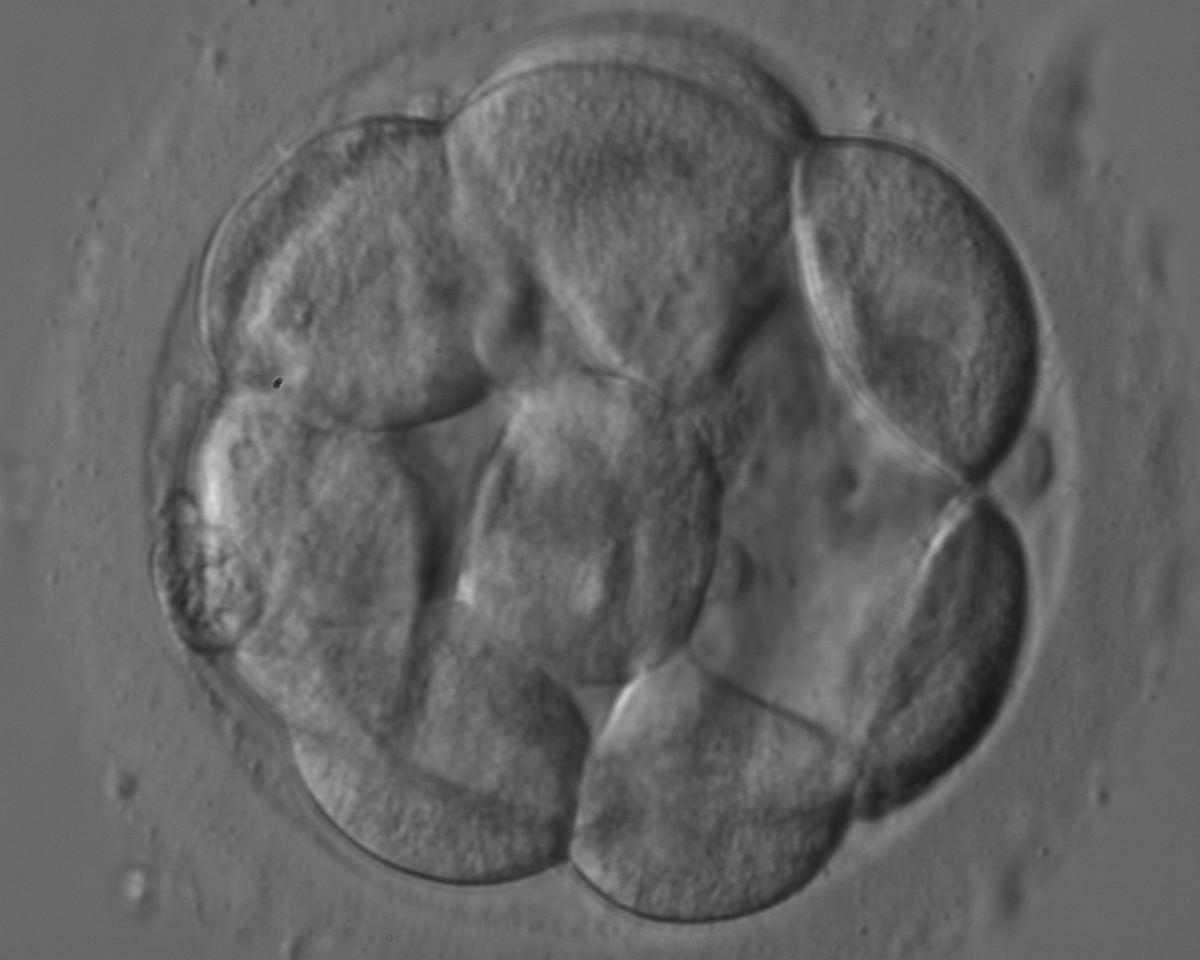
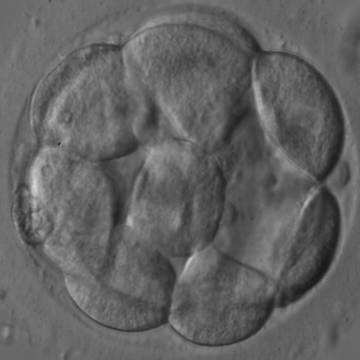
Figure 304
An early blastocyst with a cavity occupying <50% of the volume of the embryo. Note the early formation of the outer TE cells that are beginning to be flattened against the zona pellucida (ZP). The blastocyst was transferred and resulted in an ectopic pregnancy.
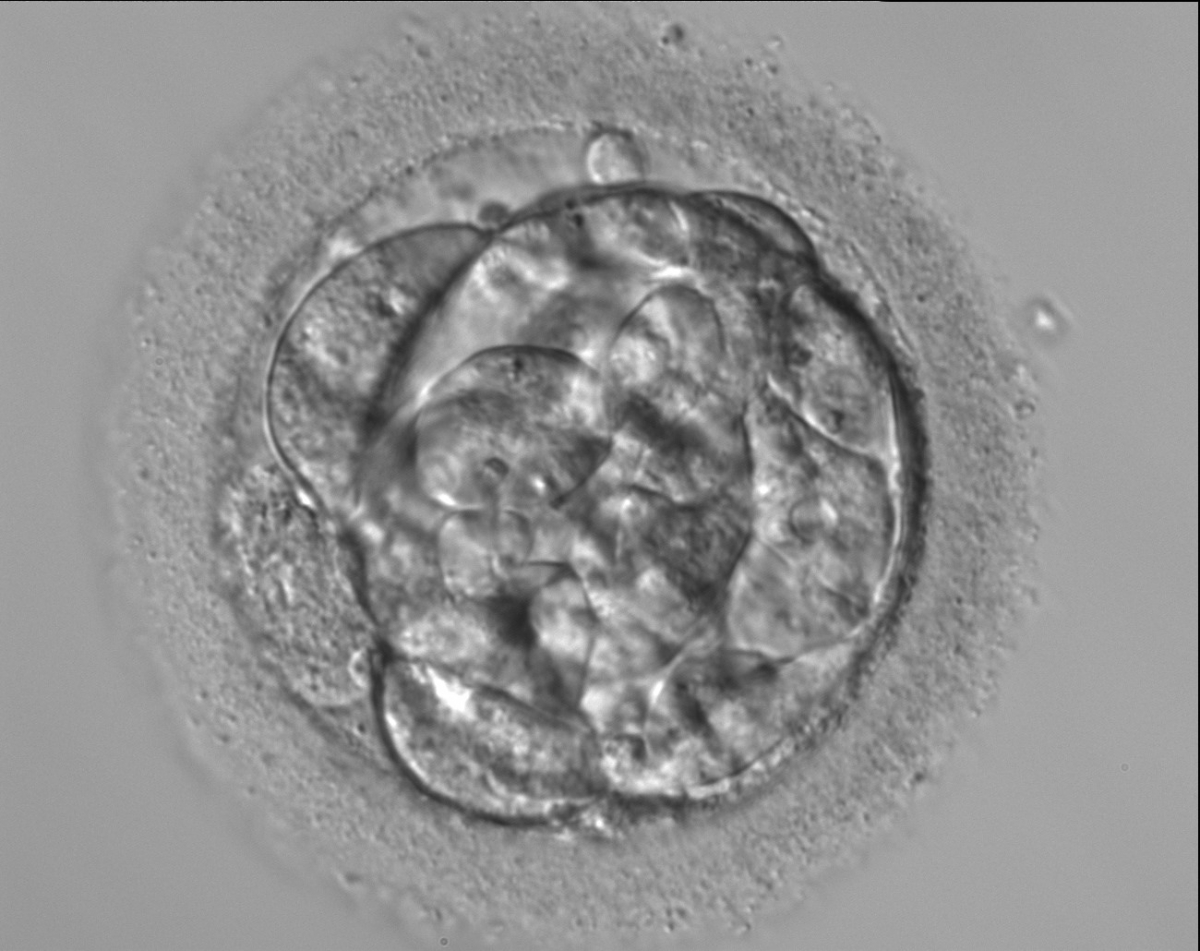
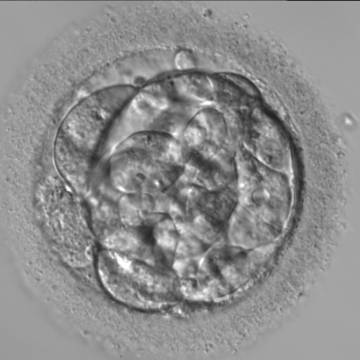
Figure 305
A very early blastocyst with a small cavity appearing centrally and can be seen most obviously at the 12 o'clock position in this view. Note the cellular debris that is not incorporated into the early blastocyst formation that has been sequestered to the perivitelline space (PVS).
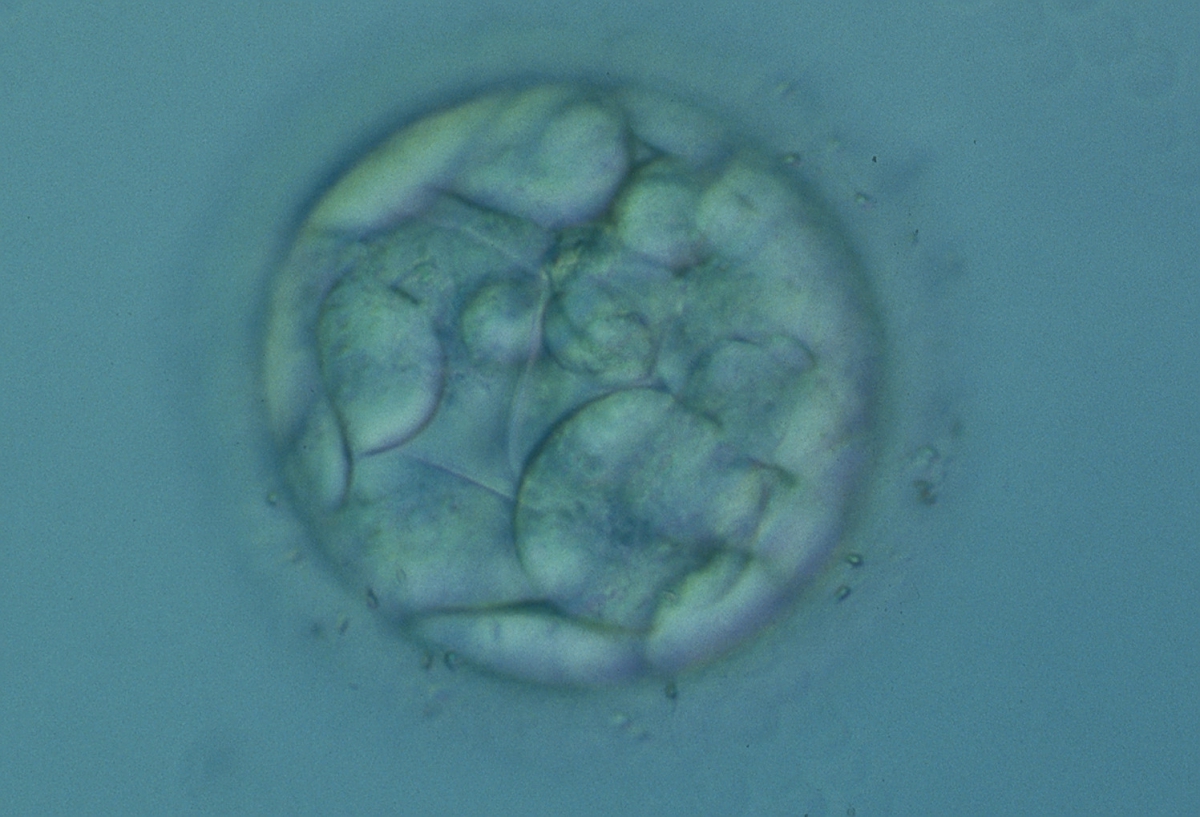
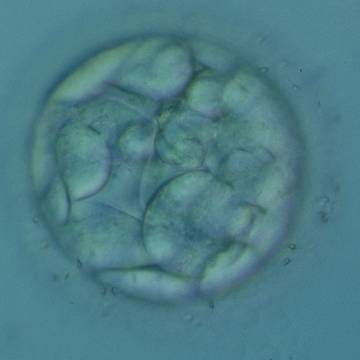
Figure 306
An early blastocyst with a cavity occupying <50% of the volume of the embryo. Note the flattened squamous-like trophectoderm (TE) cells lining the left half of the cavity in this view. The blastocyst was transferred but failed to implant.
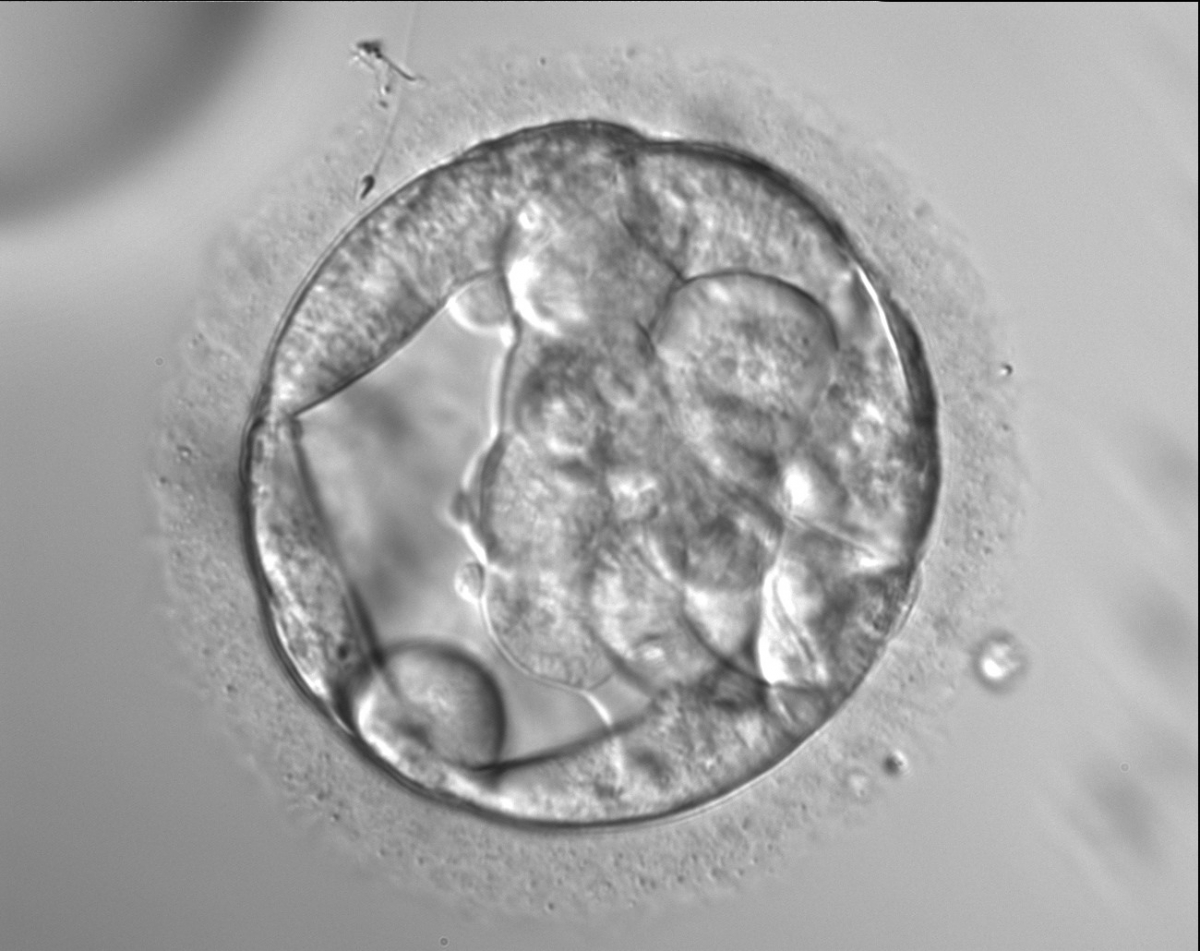
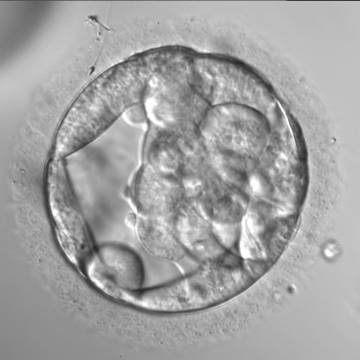
Figure 307
An early blastocyst with a cavity occupying almost 50% of the volume of the embryo. Note the large flattened TE cells lining the initial blastocoel cavity and the single spermatozoon embedded in the ZP at the 11 o'clock position in this view. The blastocyst was transferred but the outcome is unknown.
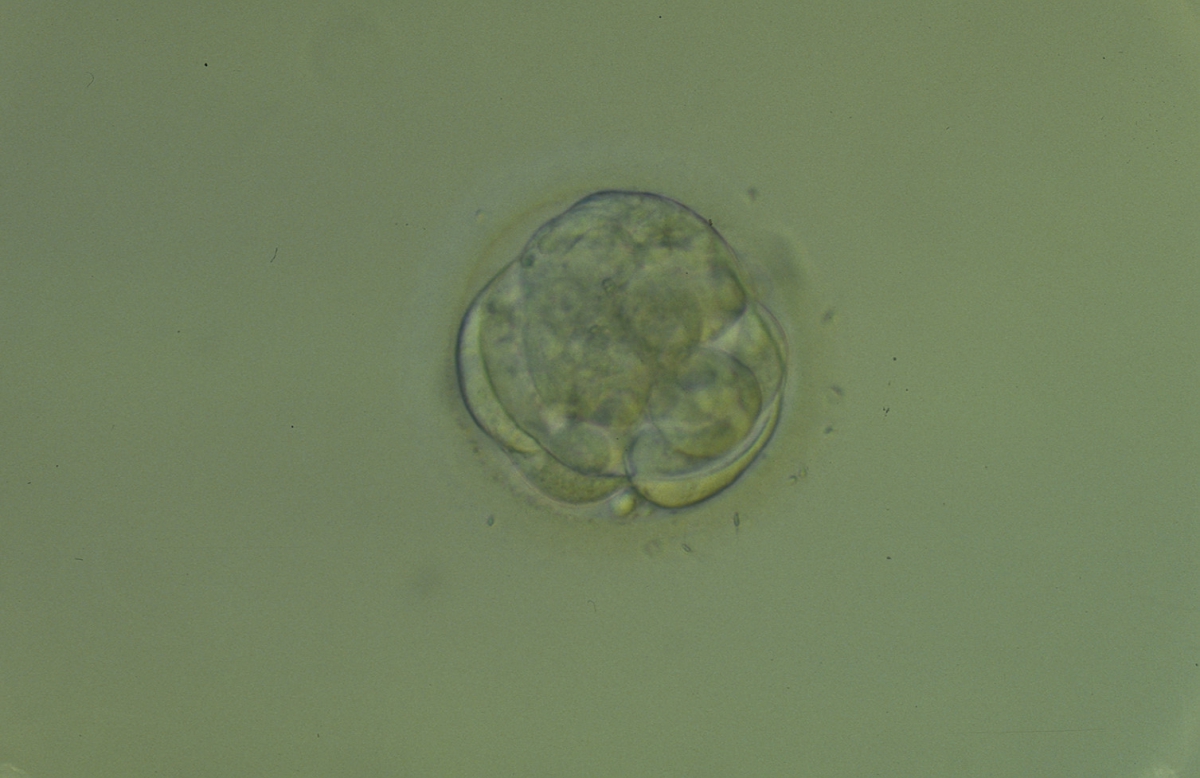
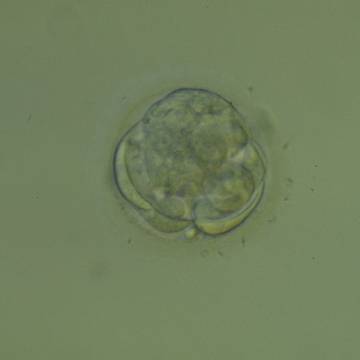
Figure 308
An early blastocyst with a cavity beginning to be formed centrally. Note the early formation of the flattened TE cells which at this stage are large, with two cells stretching to line the cavity from the 9 o'clock to the 3 o'clock position in this view. The blastocyst was transferred but failed to implant.
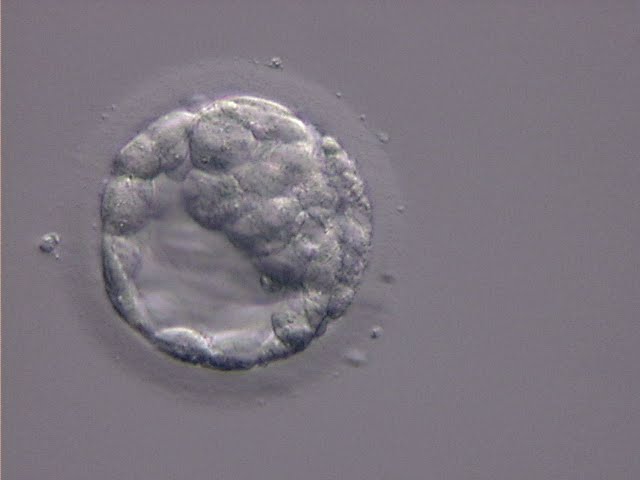
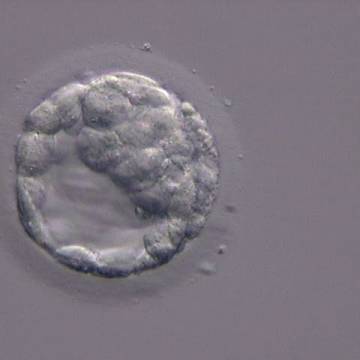
Figure 309
An early blastocyst with the cavity clearly visible and occupying half the volume of the embryo. The blastocyst was transferred but the outcome is unknown.
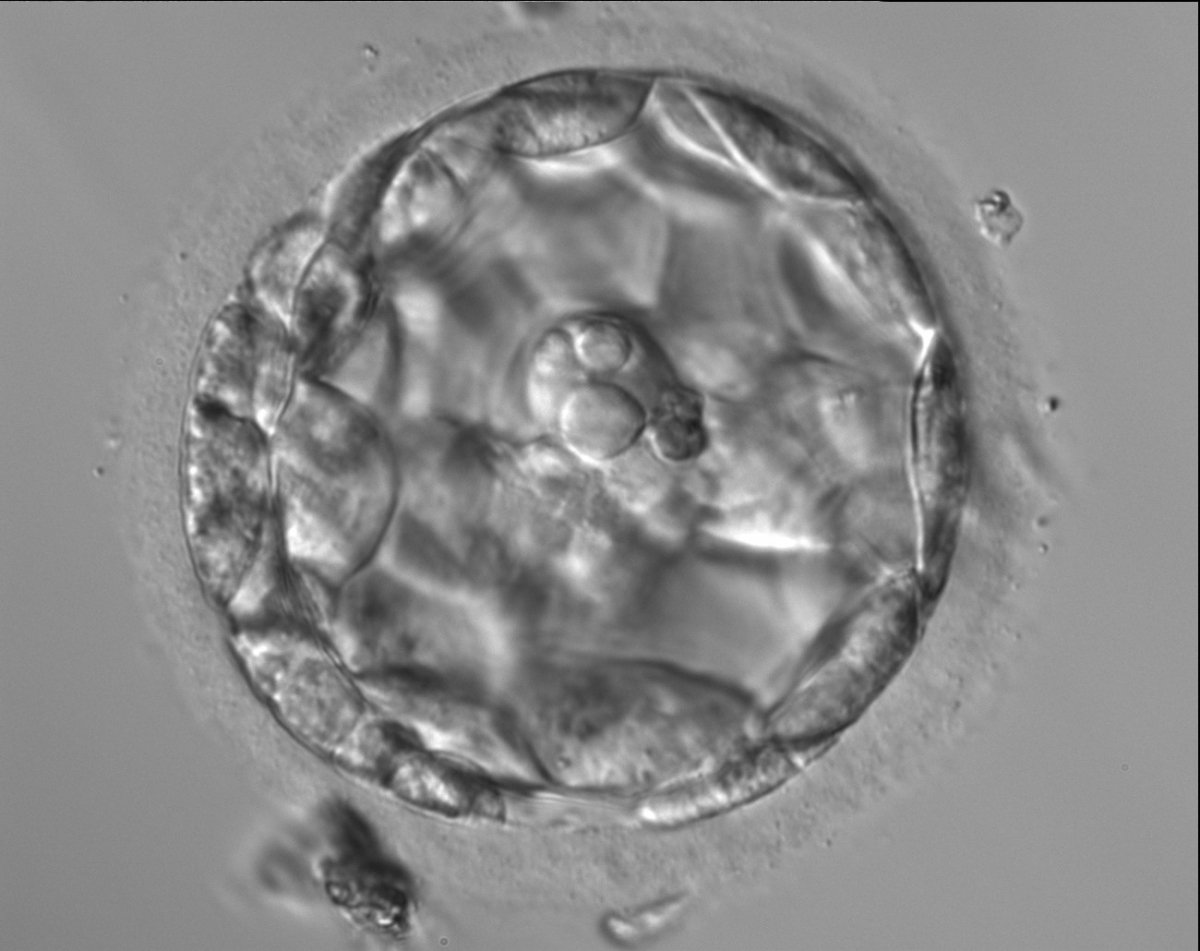
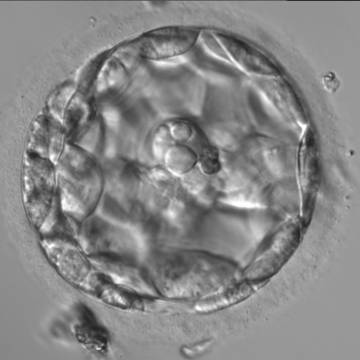
Figure 310
An early blastocyst with the cavity occupying >50% of the volume of the embryo. The overall volume of the blastocyst remains unchanged with no thinning of the ZP. The large cellular debris does not take part in the blastocyst formation.
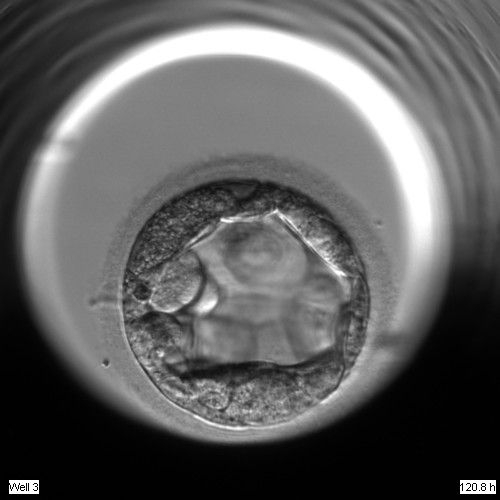
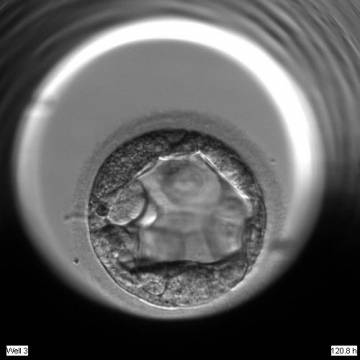
Figure 311
An early blastocyst with the cavity occupying >50% of the volume of the embryo. The TE cells are very large at this stage.
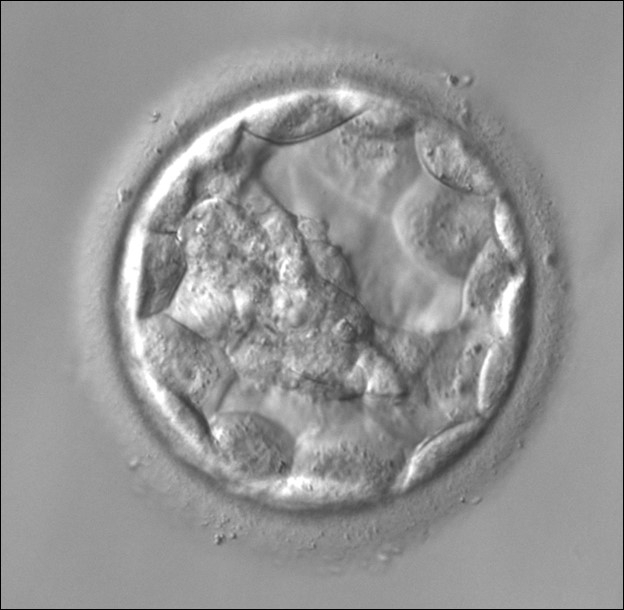
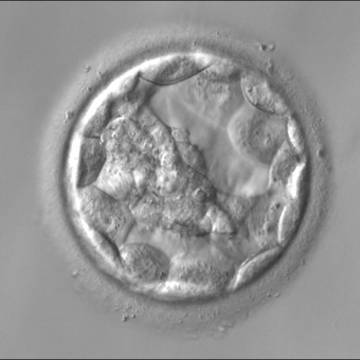
Figure 312
An early blastocyst with the cavity occupying >50% of the volume of the embryo. The overall volume of the blastocyst remains unchanged with no thinning of the ZP. The early ICM can be seen on the left half of the blastocyst in this view. The blastocyst was transferred and implanted.
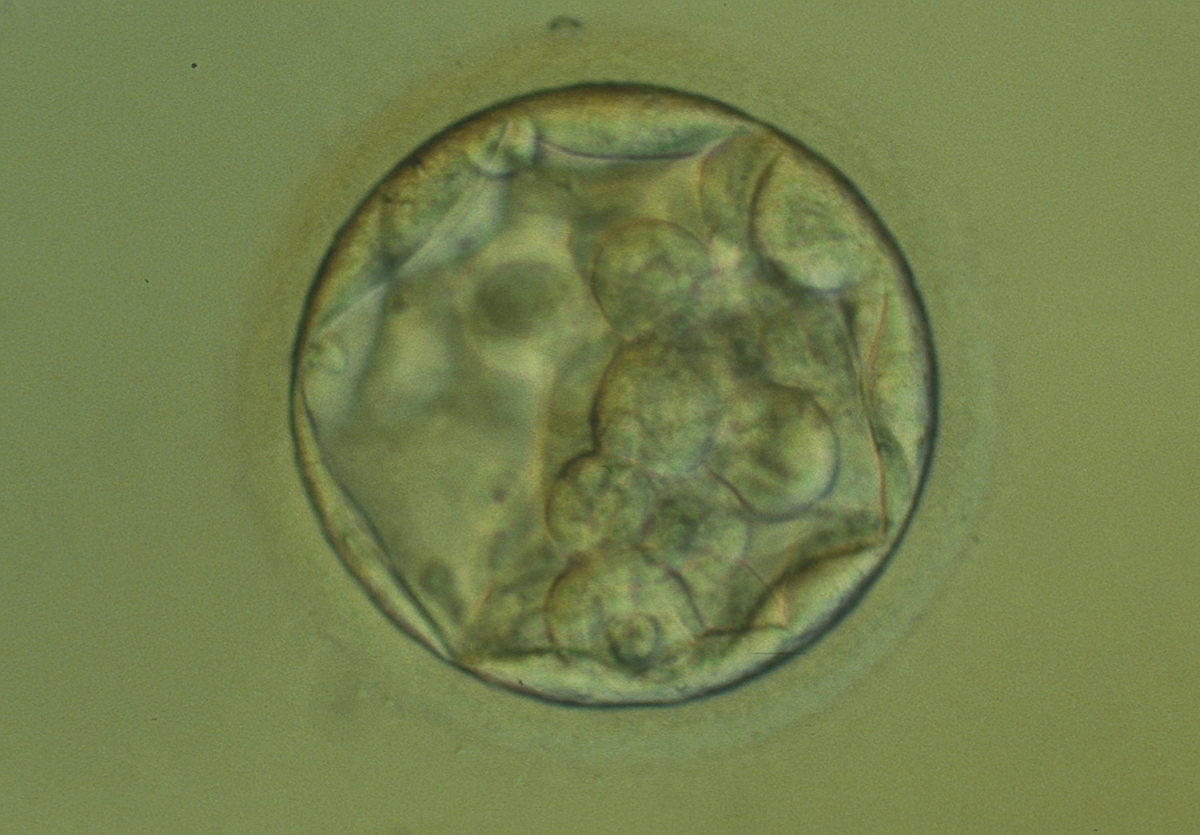
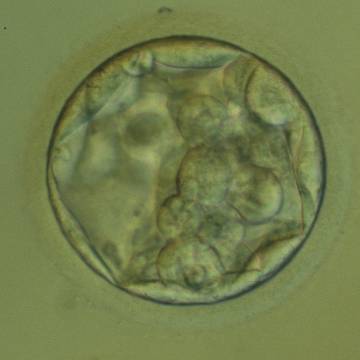
Figure 313
An early blastocyst with the cavity occupying >50% of the volume of the embryo. The overall volume of the blastocyst remains unchanged with no thinning of the ZP. The early ICM can be seen at the base of the blastocyst in this view. The blastocyst was transferred but failed to implant.
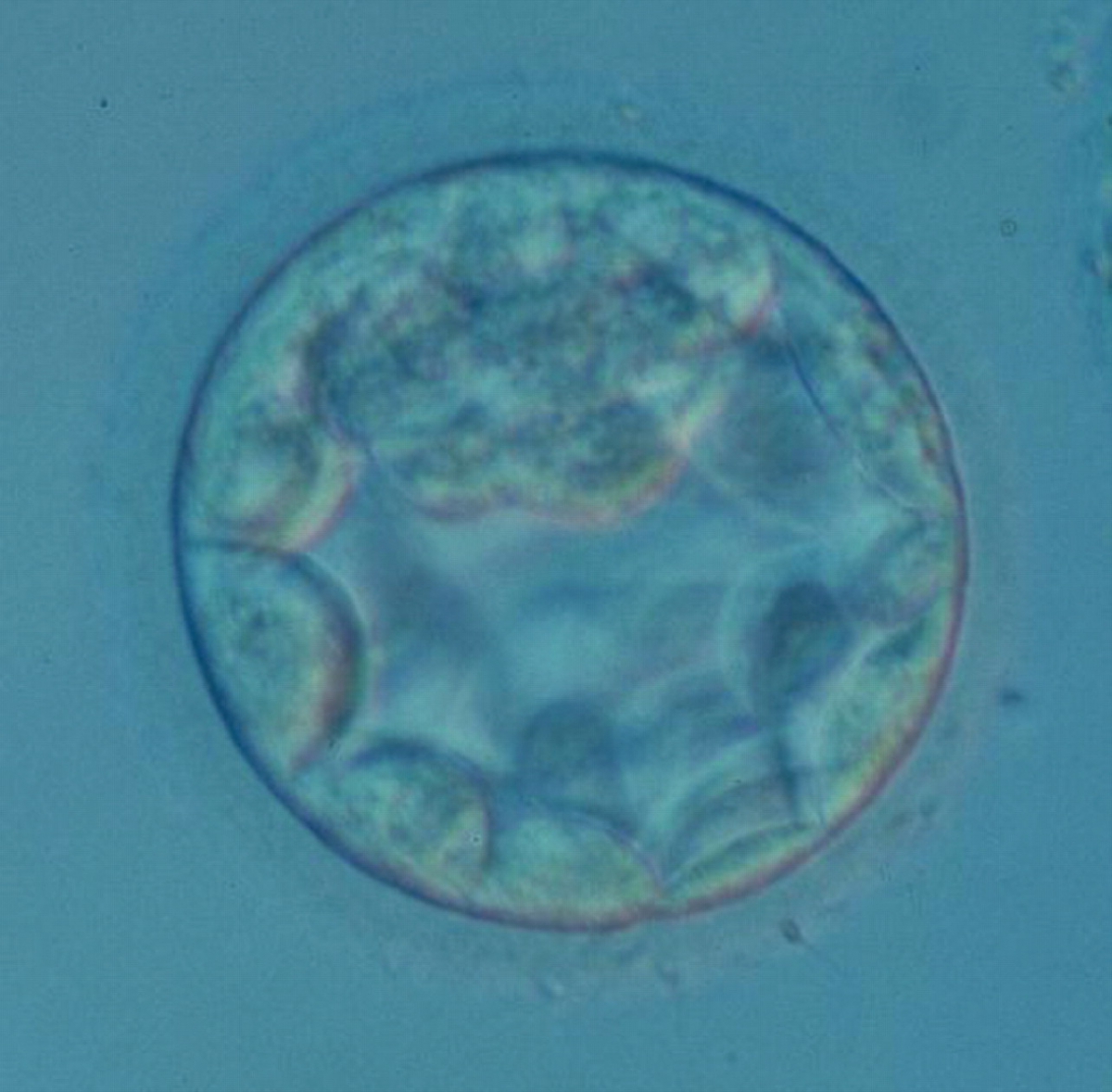
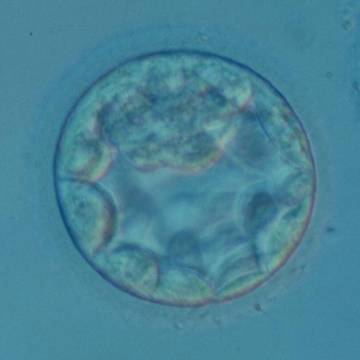
Figure 314
An early blastocyst with the cavity occupying >50% of the volume of the embryo. The overall volume of the blastocyst remains unchanged with no thinning of the ZP. The early ICM can be seen at the 12 o'clock position in this view. The blastocyst was transferred and resulted in a biochemical pregnancy.
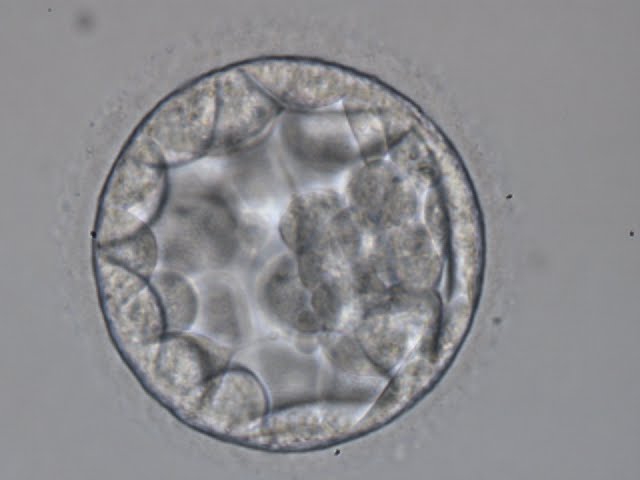
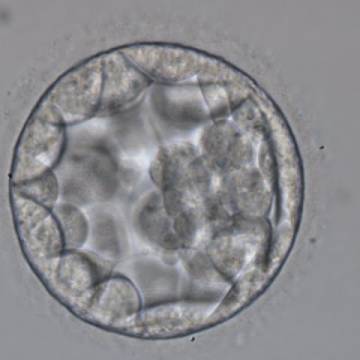
Figure 315
Blastocyst (Grade 3:2:1) showing a cavity occupying the total volume of the embryo. The ICM can be seen at the 3 o′clock position in this view and is loosely made up of only a few cells. The blastocyst was transferred but the outcome is unknown.
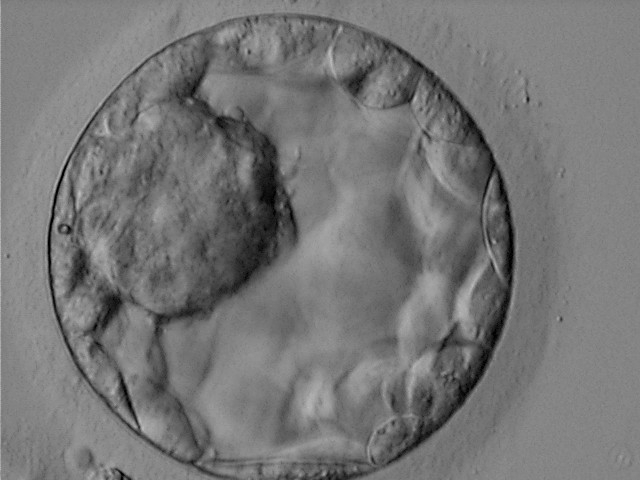
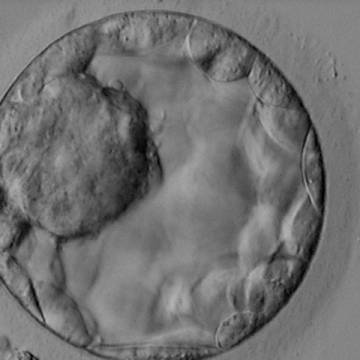
Figure 316
Blastocyst (Grade 3:1:1) showing a very large, mushroom-shaped ICM at the 10 o′clock position in this view. The ICM is made up of many cells that are tightly compacted. The blastocyst was transferred and resulted in the delivery of a healthy girl.
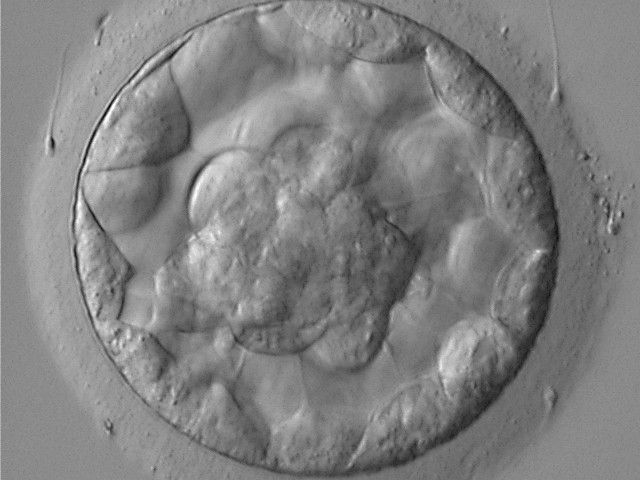
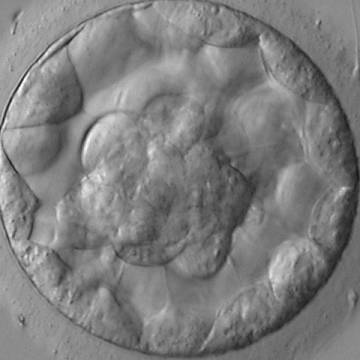
Figure 317
Blastocyst (Grade 3:1:1) showing a compact ICM at the base of the blastocyst in this view. The blastocyst was transferred and resulted in the delivery of a healthy boy.
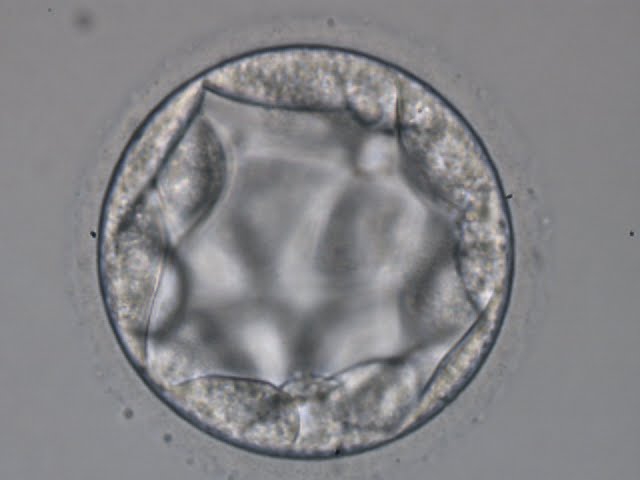
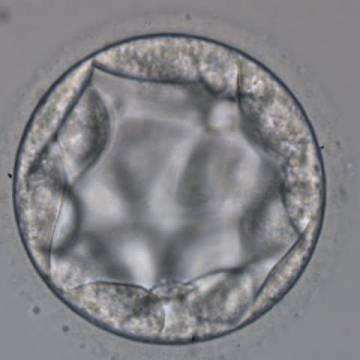
Figure 318
Blastocyst (Grade 3:3:2) with no clearly identifiable ICM and TE cells that in places are quite large and stretch over great distances to reach the next cell.
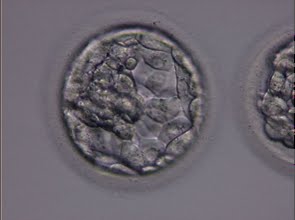
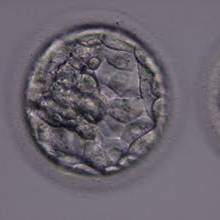
Figure 319
Blastocyst (Grade 3:1:1) with a dense ICM clearly visible at the 10 o′clock position in this view. The TE cells are variable in size but form a cohesive epithelium. The blastocyst was transferred but the outcome is unknown.
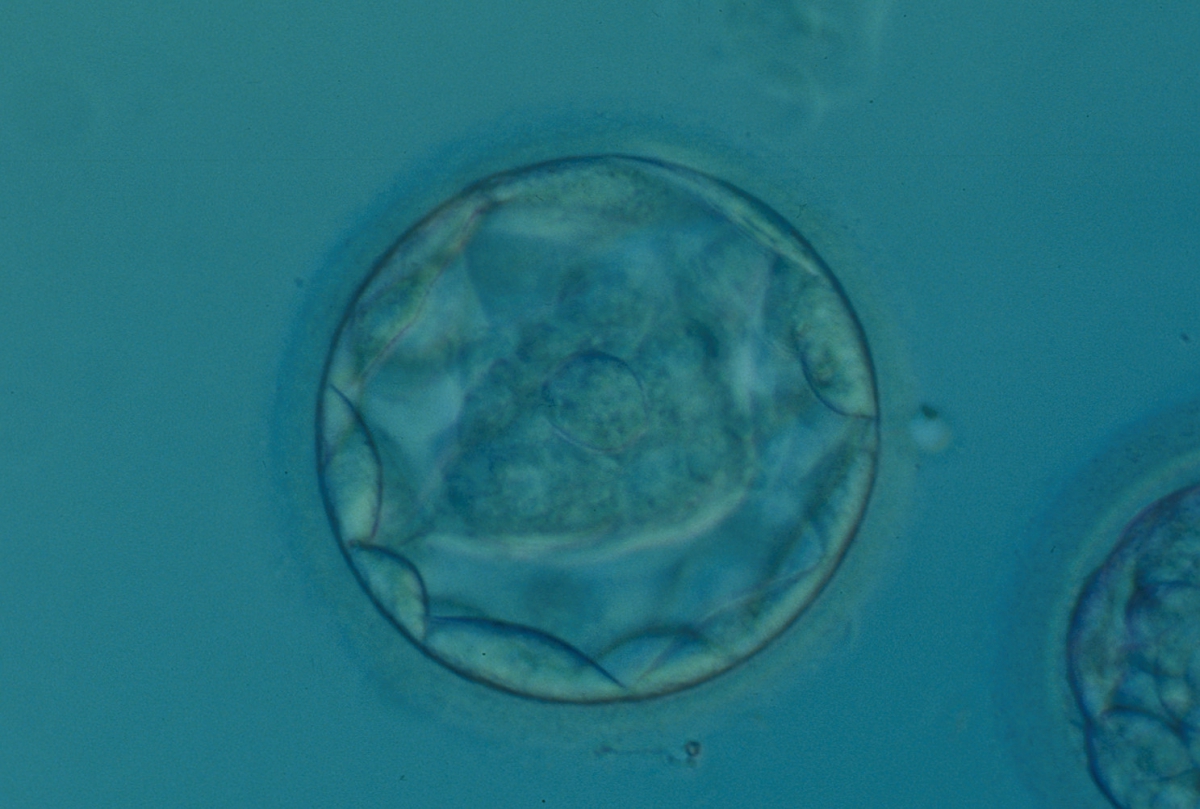
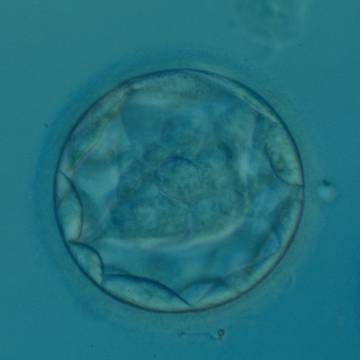
Figure 320
Blastocyst (Grade 3:1:1) with a dense, almost triangular, ICM clearly visible at the base of the blastocyst in this view. The blastocyst was transferred but failed to implant.
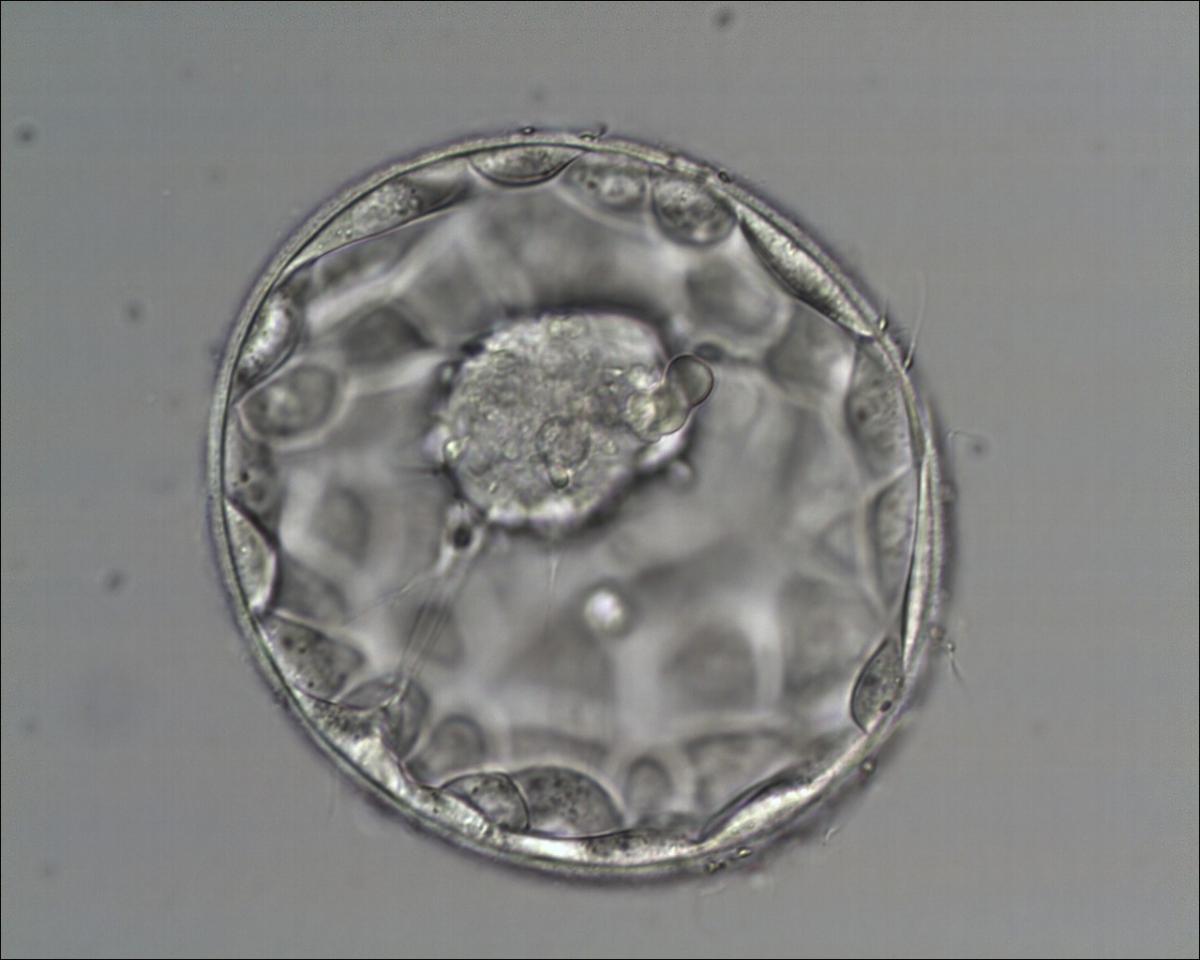
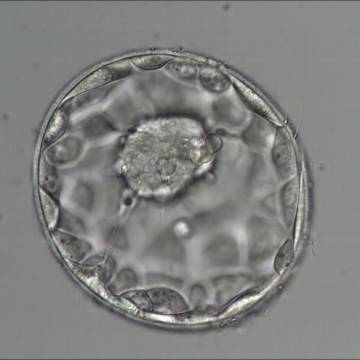
Figure 321
Good quality expanded blastocyst (Grade 4:1:1) with a large mushroom-shaped ICM. The blastocyst is now a greater volume than the original volume of the embryo and the ZP is thinned. There appears to be cytoplasmic strings extending from the ICM to the TE. The blastocyst was transferred and implanted.
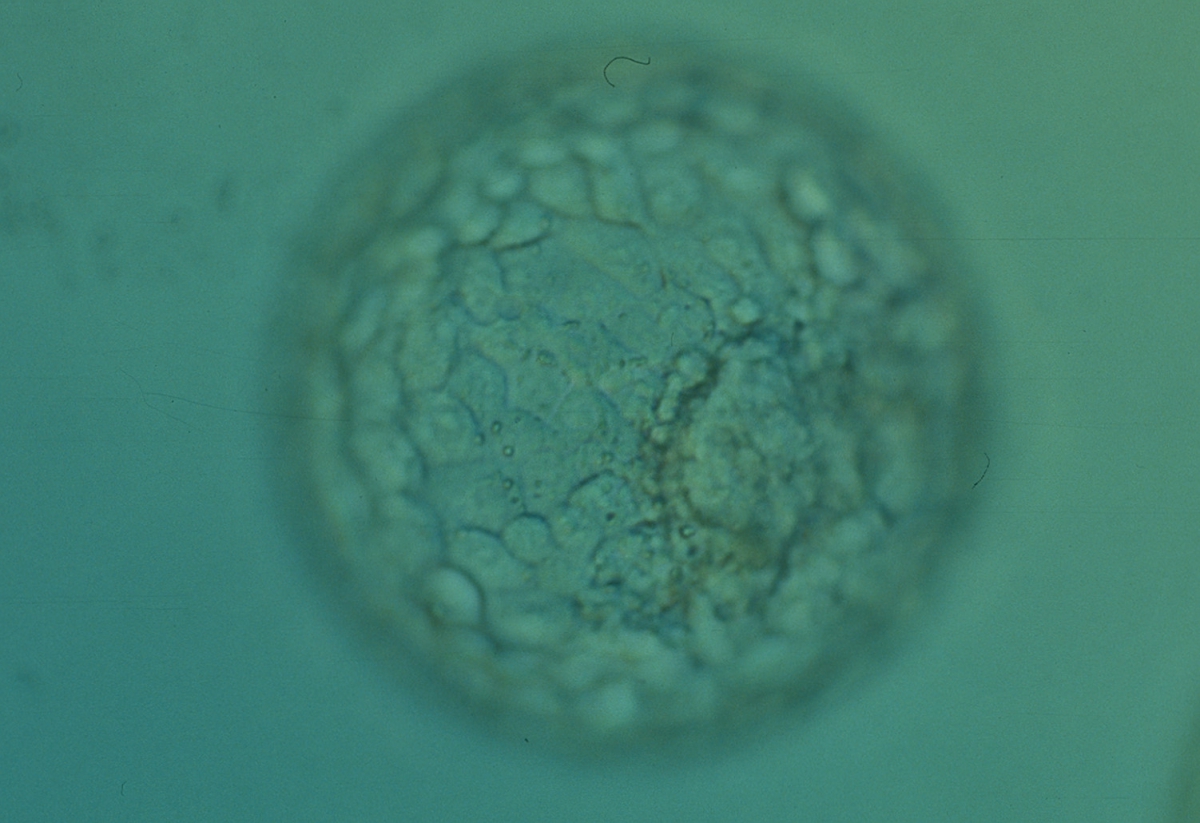
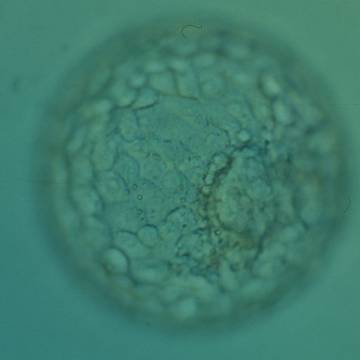
Figure 322
Expanded blastocyst (Grade 4:1:1) with an ICM clearly visible at the 4 o′clock position in this view. There are very many evenly sized cells making up a cohesive TE that surround the enlarged blastocoel cavity. The ZP is very thin. The blastocyst was transferred but the outcome is unknown.

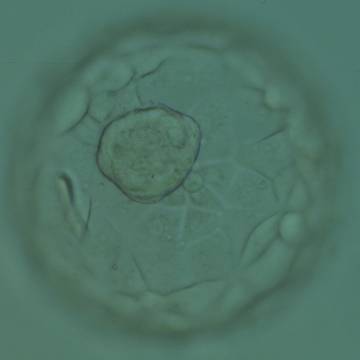
Figure 323
Expanded blastocyst (Grade 4:1:1) showing a large ICM at the base of the blastocyst in this view. The ICM is made up of many cells that are tightly compacted. The blastocyst volume is now larger than the original volume of the embryo causing the ZP to thin. The blastocyst was transferred but the outcome is unknown.
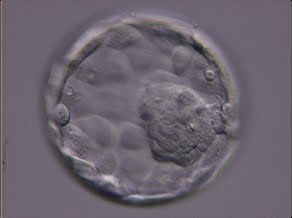
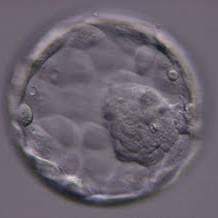
Figure 324
Expanded blastocyst (Grade 4:1:1) showing a large ICM at the 4 o′clock position in this view. The ICM is made up of many cells and is very compact. The blastocoel cavity is now larger than the original volume of the embryo and the ZP is very thin. The blastocyst was transferred but the outcome is unknown.
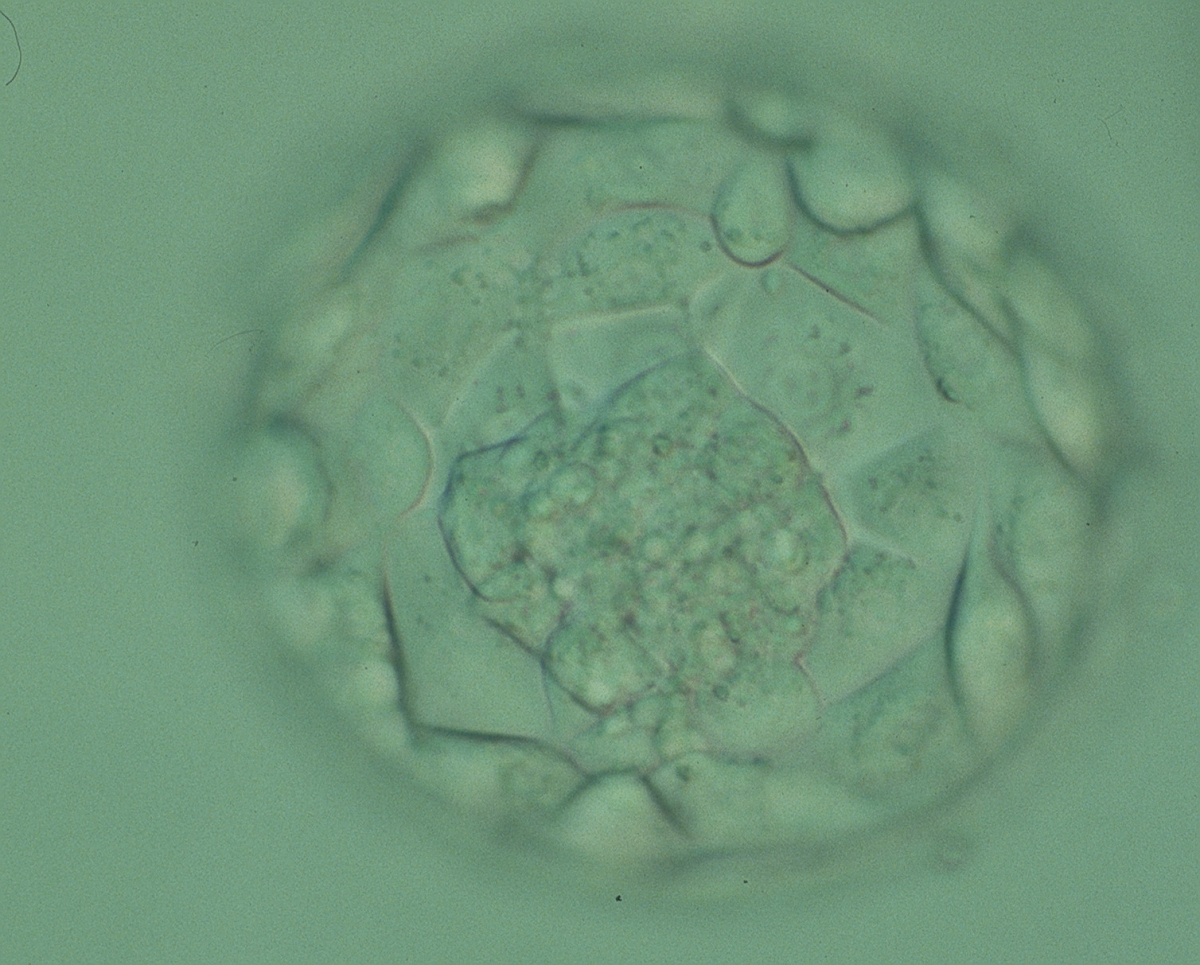
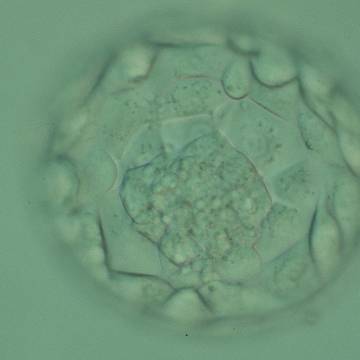
Figure 325
Expanded blastocyst (Grade 4:1:1) showing a large ICM at the base of the blastocyst in this view. There are very many TE cells forming a cohesive epithelium that lines the enlarged blastocoel cavity. The ZP is very thin. The blastocyst was transferred but failed to implant.
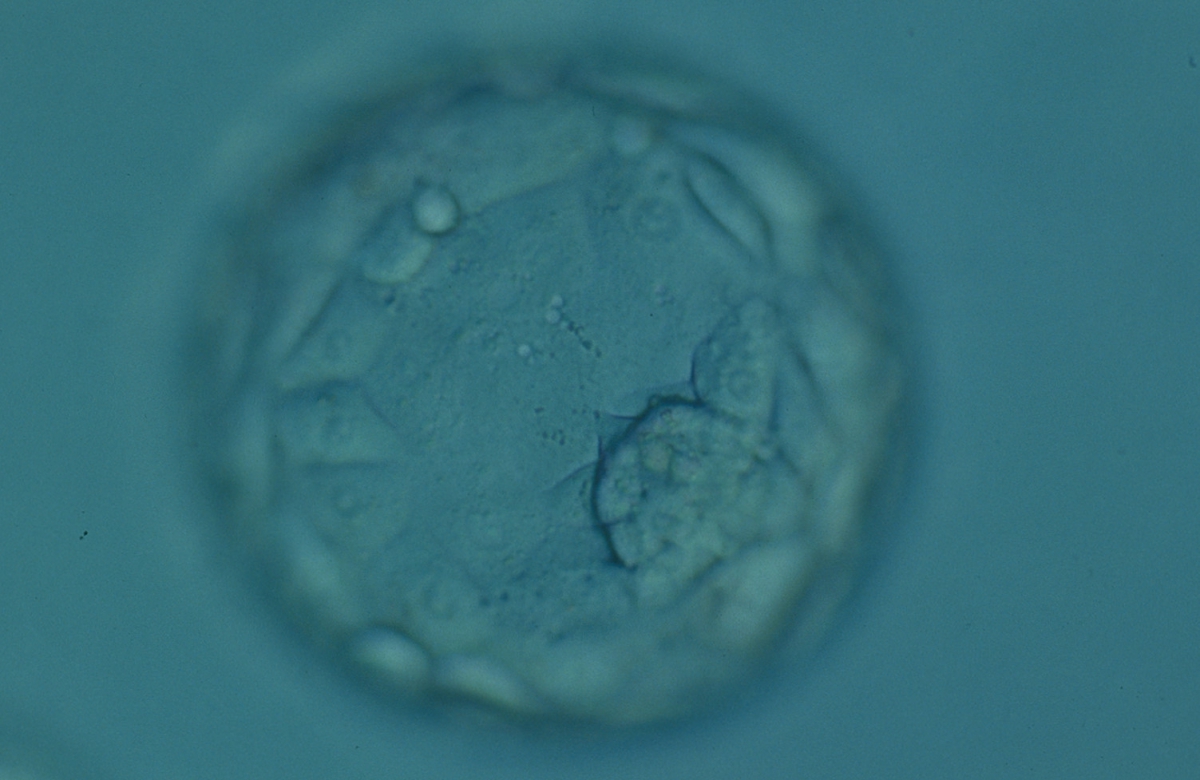
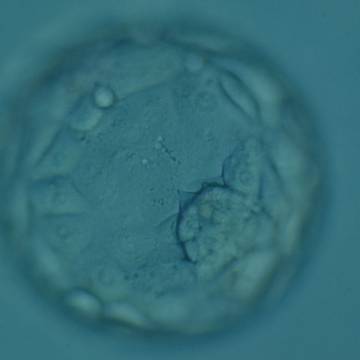
Figure 326
Expanded blastocyst (Grade 4:1:1) showing a compact ICM at the 4 o'clock position in this view. The blastocyst is now larger in volume than the original volume of the embryo causing the ZP to thin. The blastocyst was transferred but failed to implant.
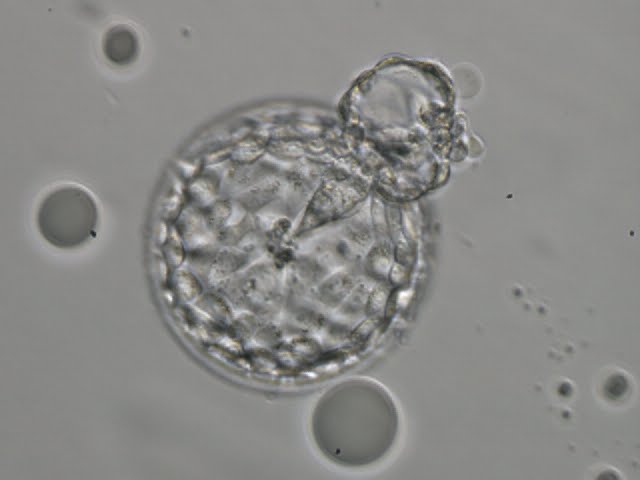
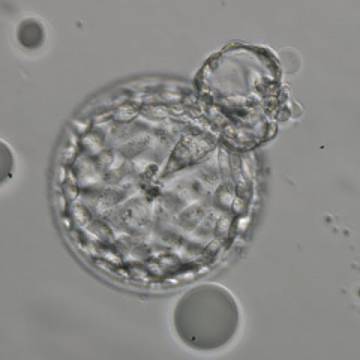
Figure 327
Hatching blastocyst (Grade 5:2:1) showing a small triangular ICM being drawn out along with the herniating TE cells at the 1 o'clock position in this view. There are very many TE cells of similar size lining the blastocoel cavity and the ZP is thinned. The blastocyst was transferred but the outcome is unknown.
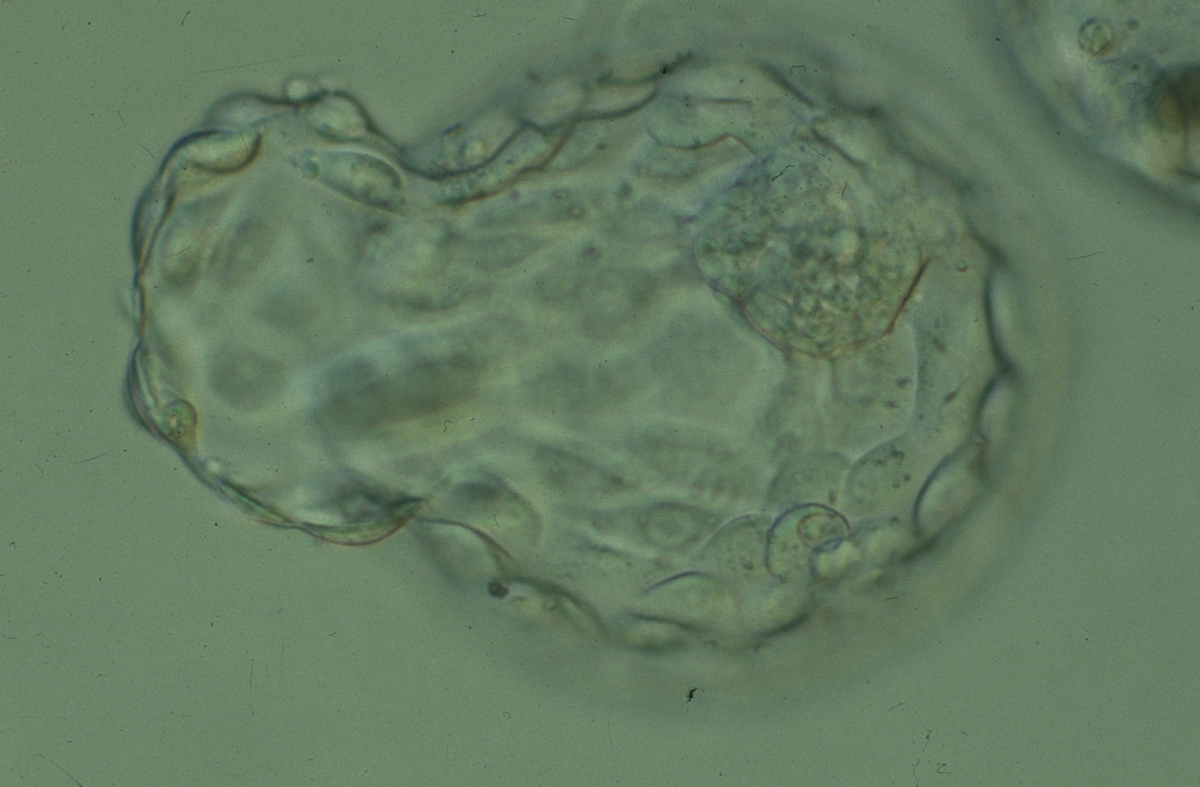
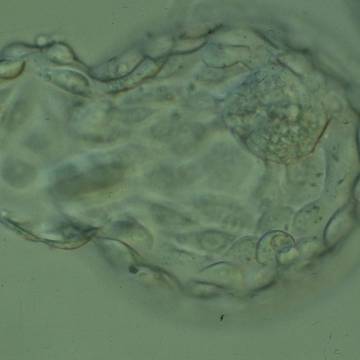
Figure 328
Hatching blastocyst (Grade 5:1:1) showing a large, compact ICM at the 1 o'clock position in this view. Approximately 25% of the blastocyst has herniated from a breach in the thinned ZP at the 8–10 o'clock positions in this view. The blastocyst was transferred but the outcome is unknown.
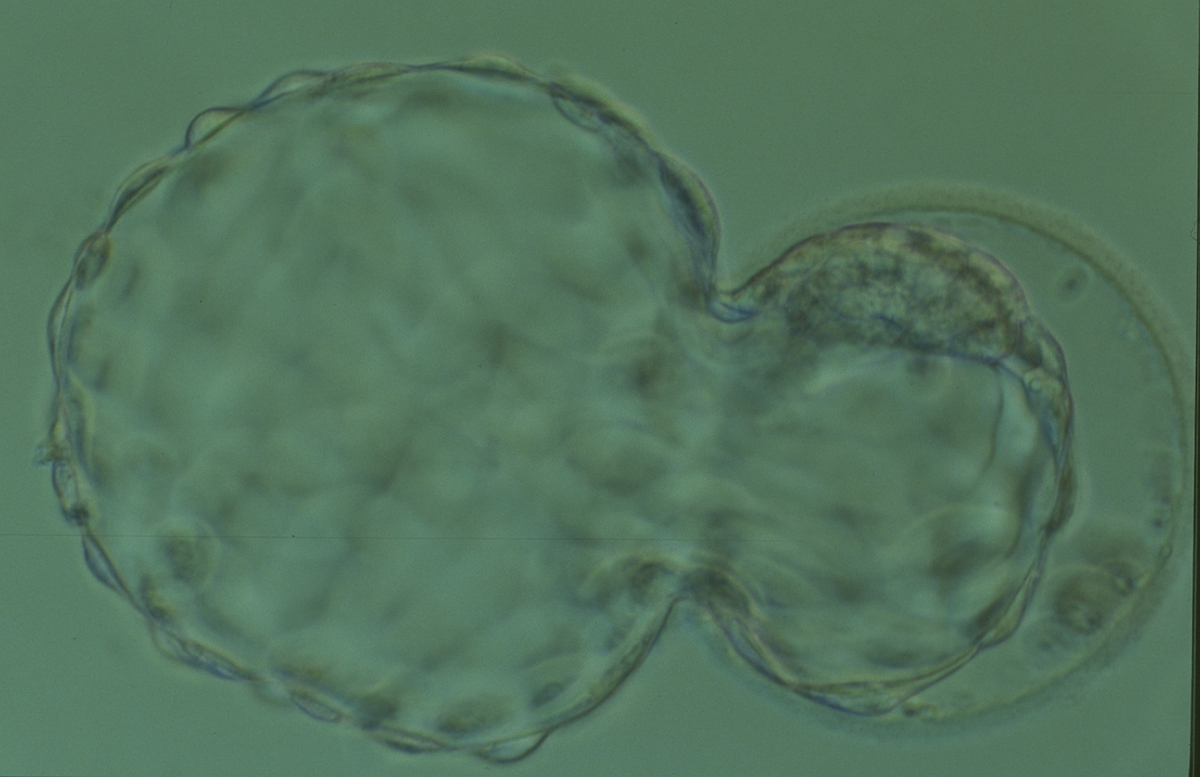
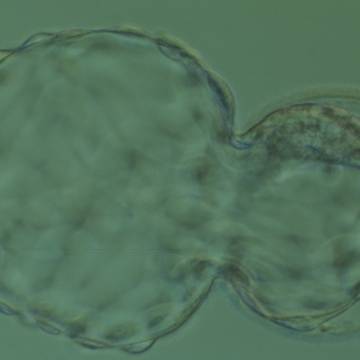
Figure 329
Hatching blastocyst (Grade 5:1:1) showing a large, compact, crescent-shaped ICM retained within the ZP at the 12 o'clock position in this view. There are very many TE cells and almost 75% of the blastocyst has herniated out through a breach in the ZP at the 8–10 o'clock positions in this view. The blastocyst was transferred but the outcome is unknown.
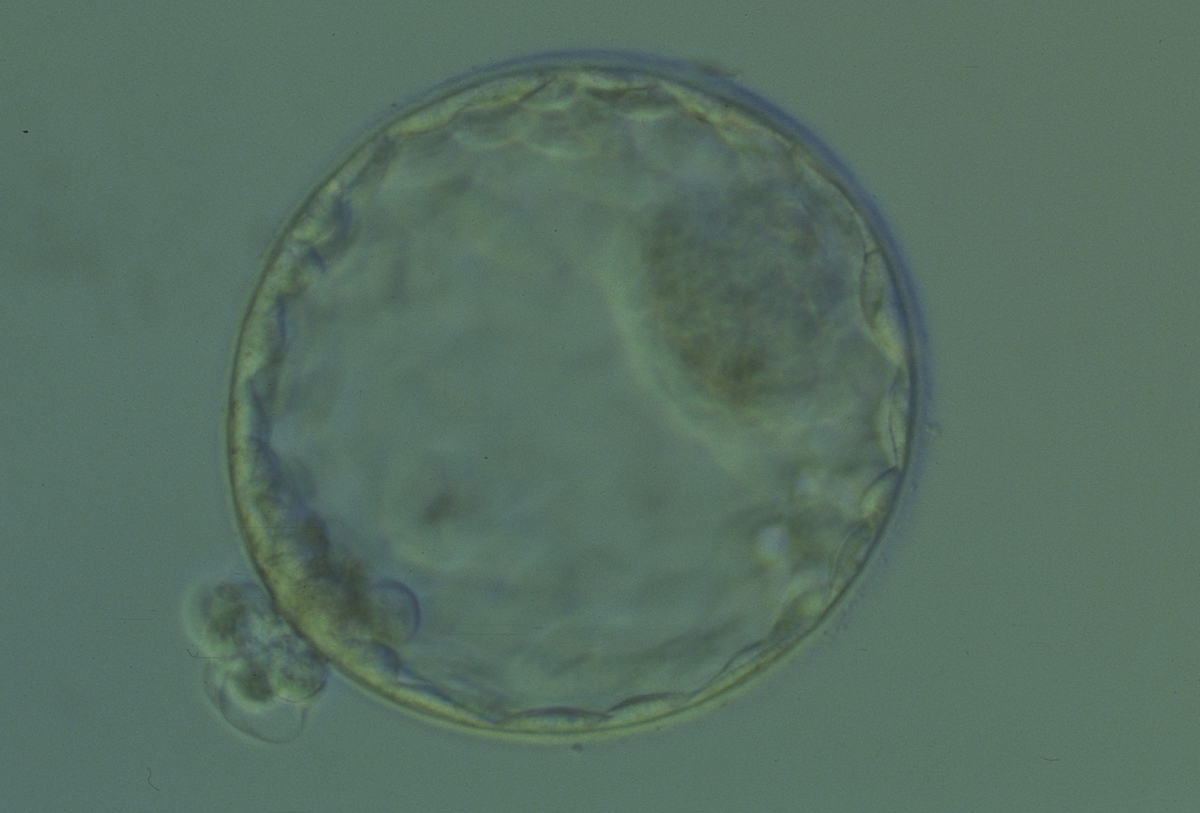
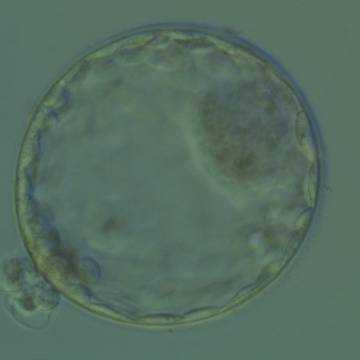
Figure 330
Hatching blastocyst (Grade 5:1:1) showing a large, compact ICM at the 2 o'clock position in this view. There are many TE cells of equivalent size lining the blastocoel cavity and several TE cells are herniating through a breach in the thinned ZP at the 8 o'clock position in this view. The blastocyst was transferred and implanted.

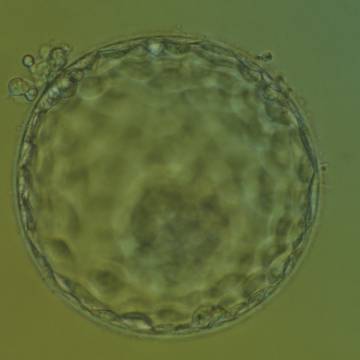
Figure 331
Hatching blastocyst (Grade 5:1:1) showing a large, compact ICM at the base of the blastocyst toward the 5 o'clock position and slightly out of focus in this view. There are very many TE cells of equivalent size making up a cohesive epithelium. Several TE cells have herniated out through a breach in the ZP at the 11 o'clock position in this view. The blastocyst was transferred and implanted.
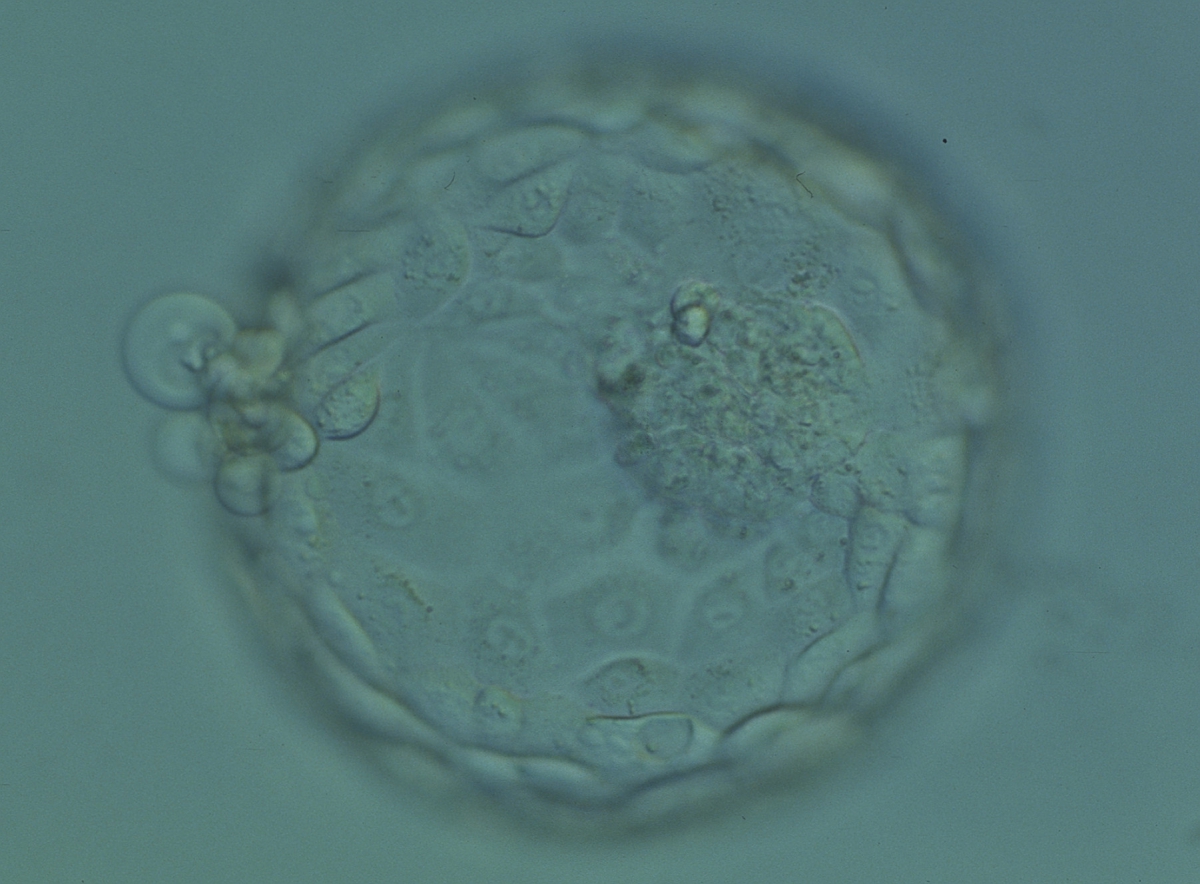
Figure 332
Hatching blastocyst (Grade 5:1:1) showing a large, compact and slightly granular ICM toward the 3 o'clock position in this view. There are very many TE cells of varying sizes making up a cohesive epithelium with several cells herniating out through a breach in the ZP at the 9 o'clock position in this view. The blastocyst was transferred but the outcome is unknown.
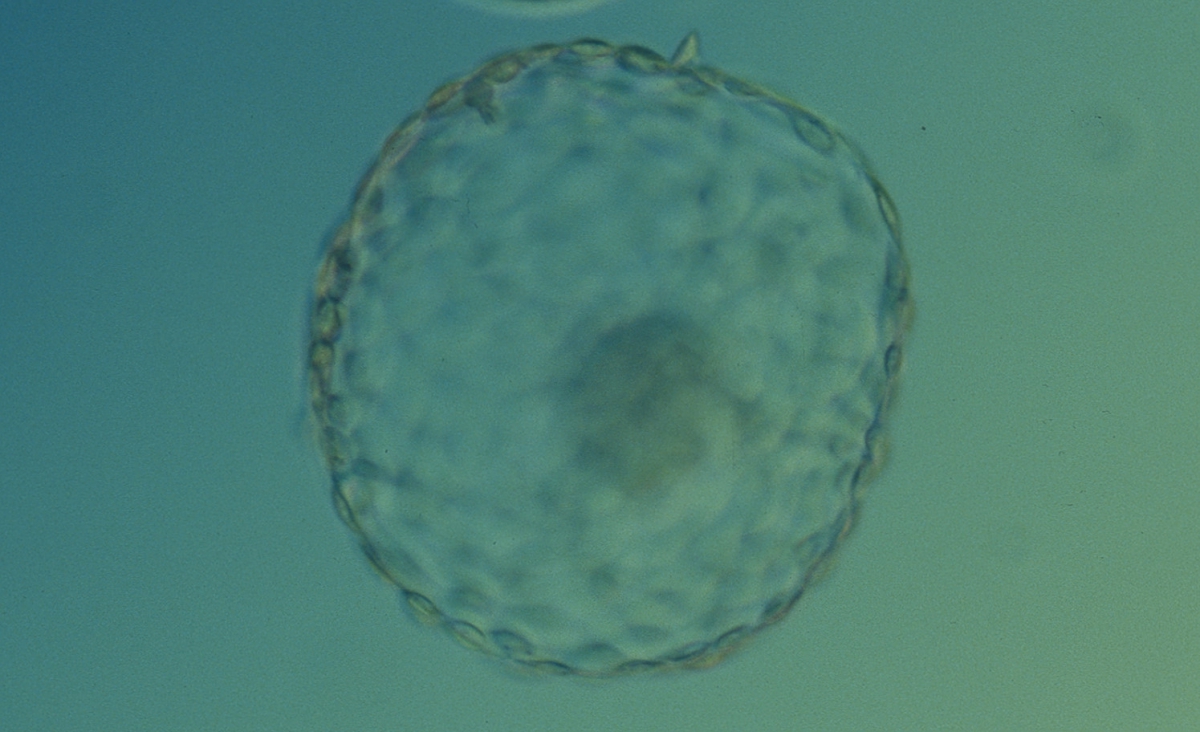
Figure 333
Hatched blastocyst (Grade 6:1:1) that is now completely free of the ZP. There is a large, compact ICM slightly out of focus at the base of the blastocyst toward the 4 o'clock position in this view. The blastocyst is now more than twice the size of the original expanded blastocyst. The blastocyst was transferred but failed to implant.
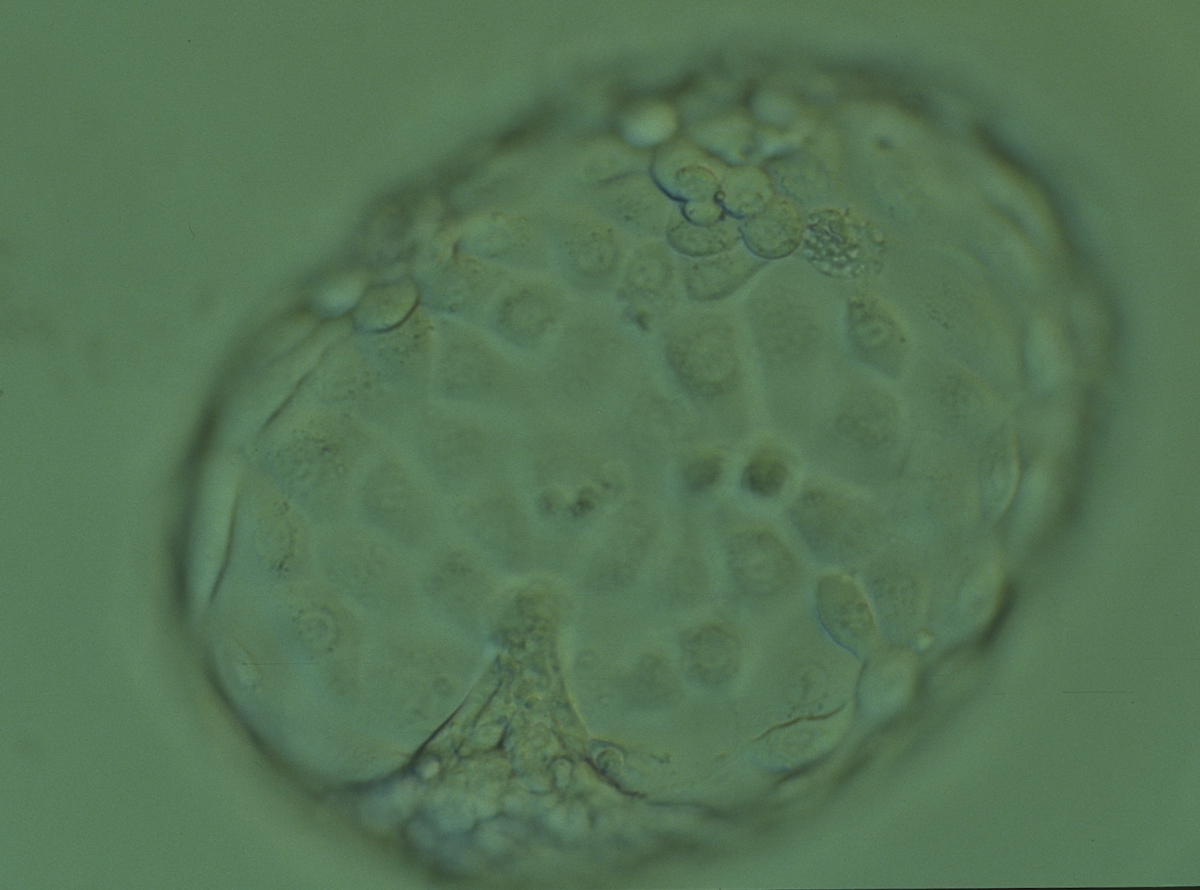
Figure 334
Hatched blastocyst (Grade 6:1:1) that is now completely free of the ZP showing a compact ICM at the 6 o'clock position in this view. The ICM appears to be connected or anchored to the TE by a broad triangular bridge. The TE cells vary in size but form a cohesive epithelium. The blastocyst is now more than twice the size of the original expanded blastocyst. The blastocyst was transferred but the outcome is unknown.
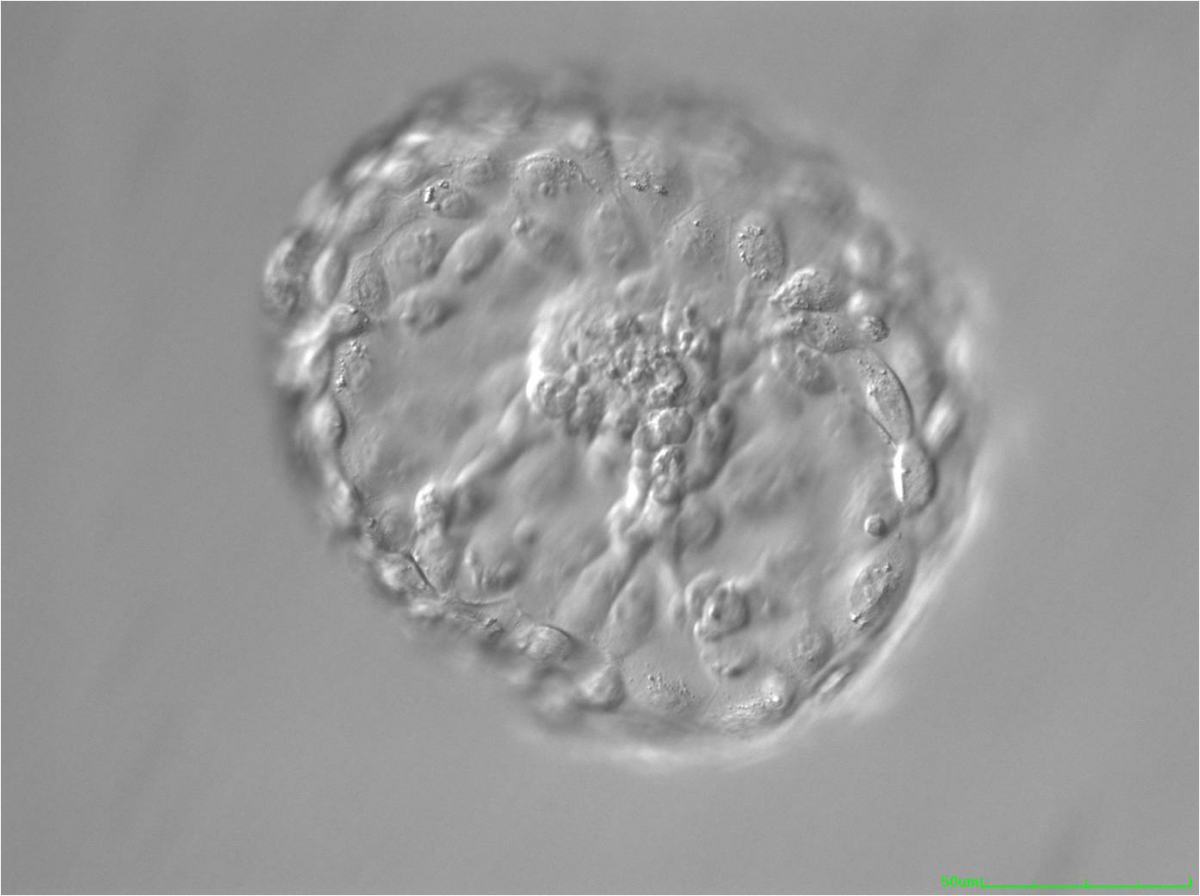
Figure 335
Hatched blastocyst (Grade 6:1:1) that is now completely free of the ZP. The ICM positioned centrally at the base of the blastocyst appears to be connected or anchored to the TE by several bridges. There are very many TE cells of similar size forming a cohesive epithelium. The blastocyst is now more than twice the size of the original expanded blastocyst.
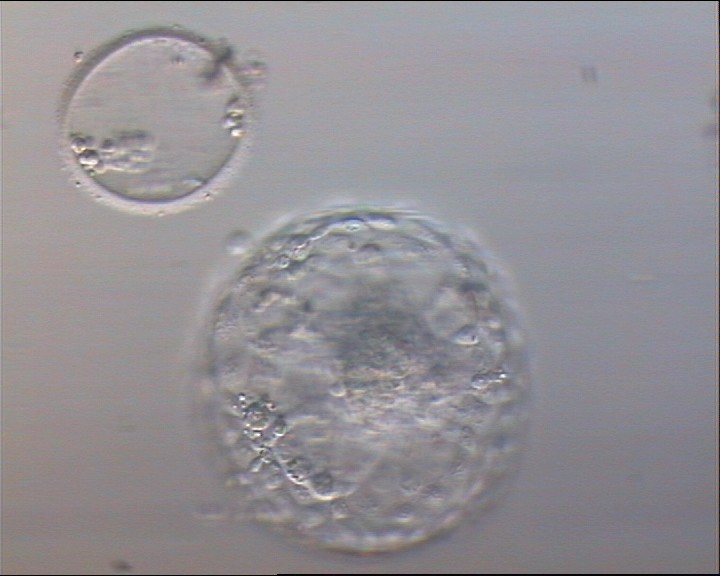
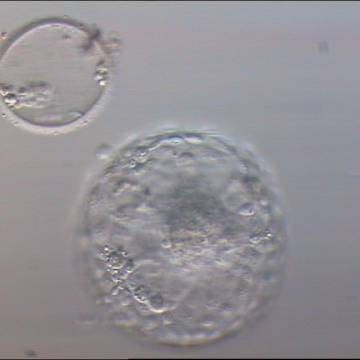
Figure 336
Hatched blastocyst (Grade 6:1:1) that is now completely free of the ZP which can be seen in the same view. Some cellular debris remains behind in the empty ZP. The ICM is large and compact at the base of the blastocyst and there are many TE cells making up a cohesive epithelium. In this view, it is possible to clearly see the increase in diameter of the blastocyst from the diameter of the original embryo which was accommodated within the ZP.
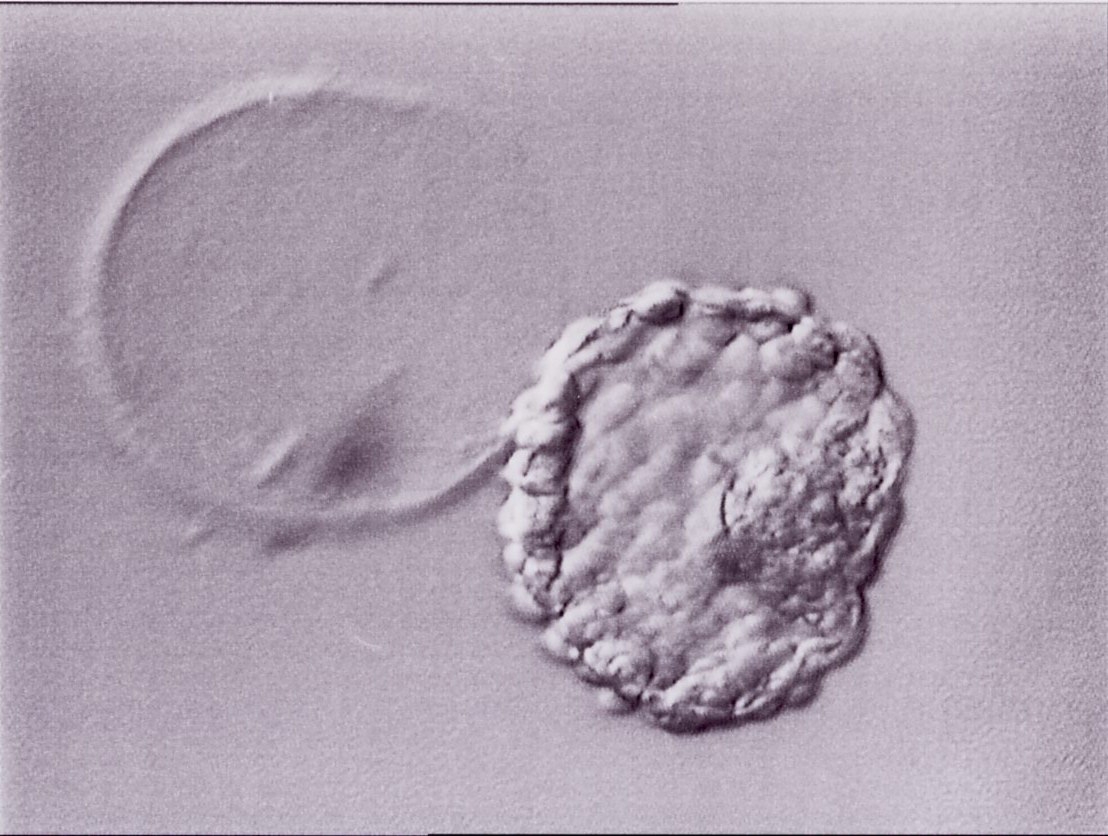
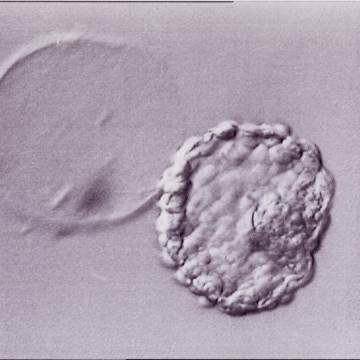
Figure 337
Hatched blastocyst (Grade 6:1:1) that is now completely free of the ZP which can be seen in the same view. The breach in the ZP is large. The ICM and TE both have many cells and the blastocyst has collapsed slightly and appears more dense. The blastocyst was transferred but the outcome is unknown.
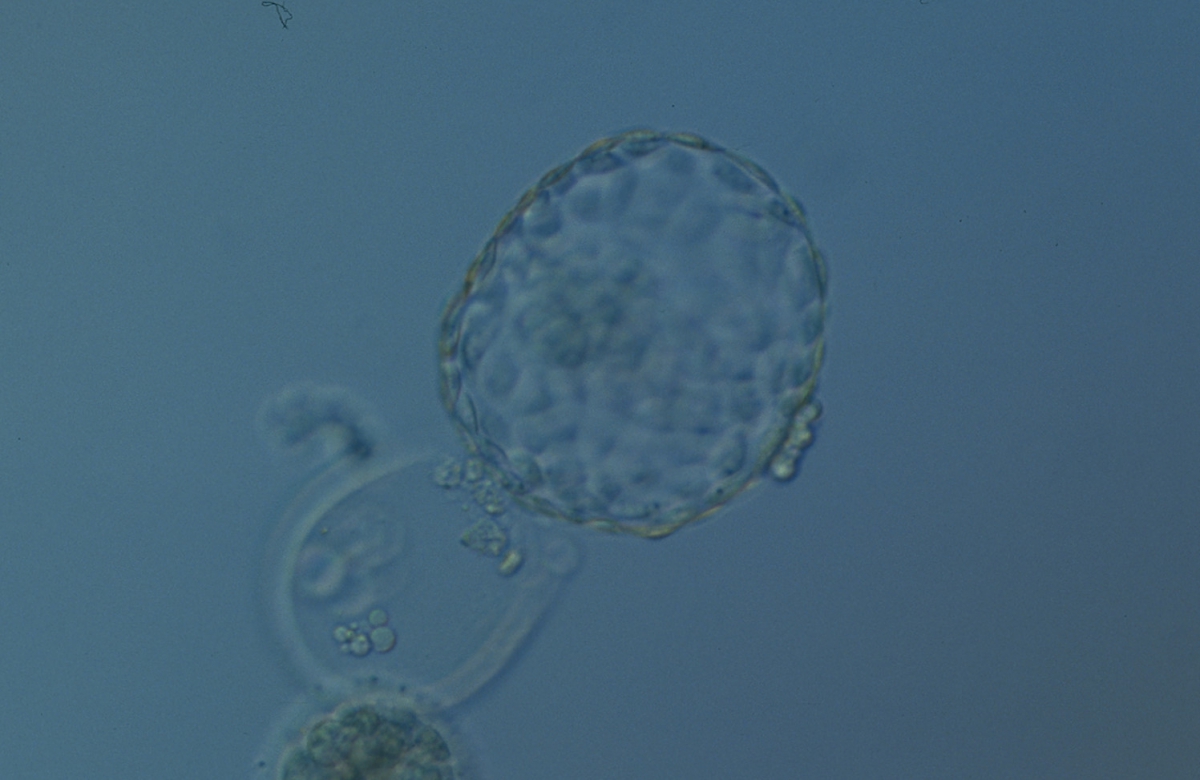
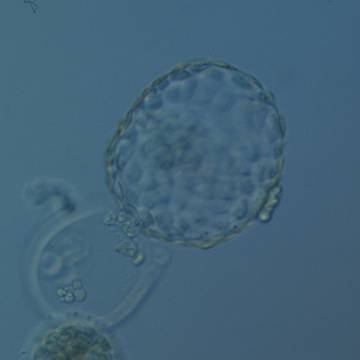
Figure 338
Hatched blastocyst (Grade 6:1:1) that is now only just free of the ZP which can also be seen in the low magnification view. The ICM is slightly out of focus in this view and is made up of many cells. The TE is similarly made up of many cells forming a cohesive epithelium. There is some cellular debris discarded in the ZP. The blastocyst was transferred but the outcome is unknown.
It is not uncommon to observe blastocysts that have collapsed or are in the process of collapsing (Figs 339–341). In this instance it is difficult to accurately grade the blastocysts and the consensus document (Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology, 2011) suggests that 1–2h is allowed to elapse before the blastocyst is re-assessed as regular cycles of expansion and collapse are normal and can be observed even without intervention as has been recorded using continuous time-lapse recording within the incubator.
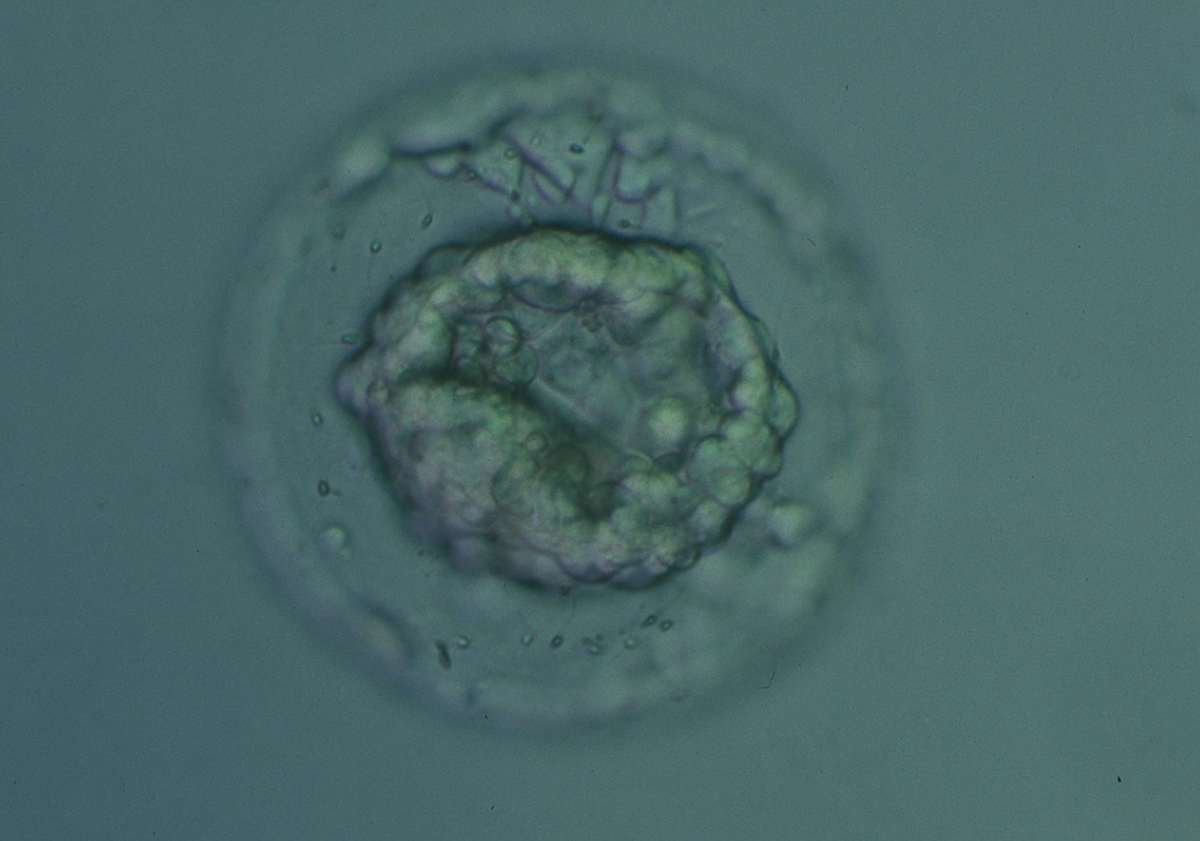
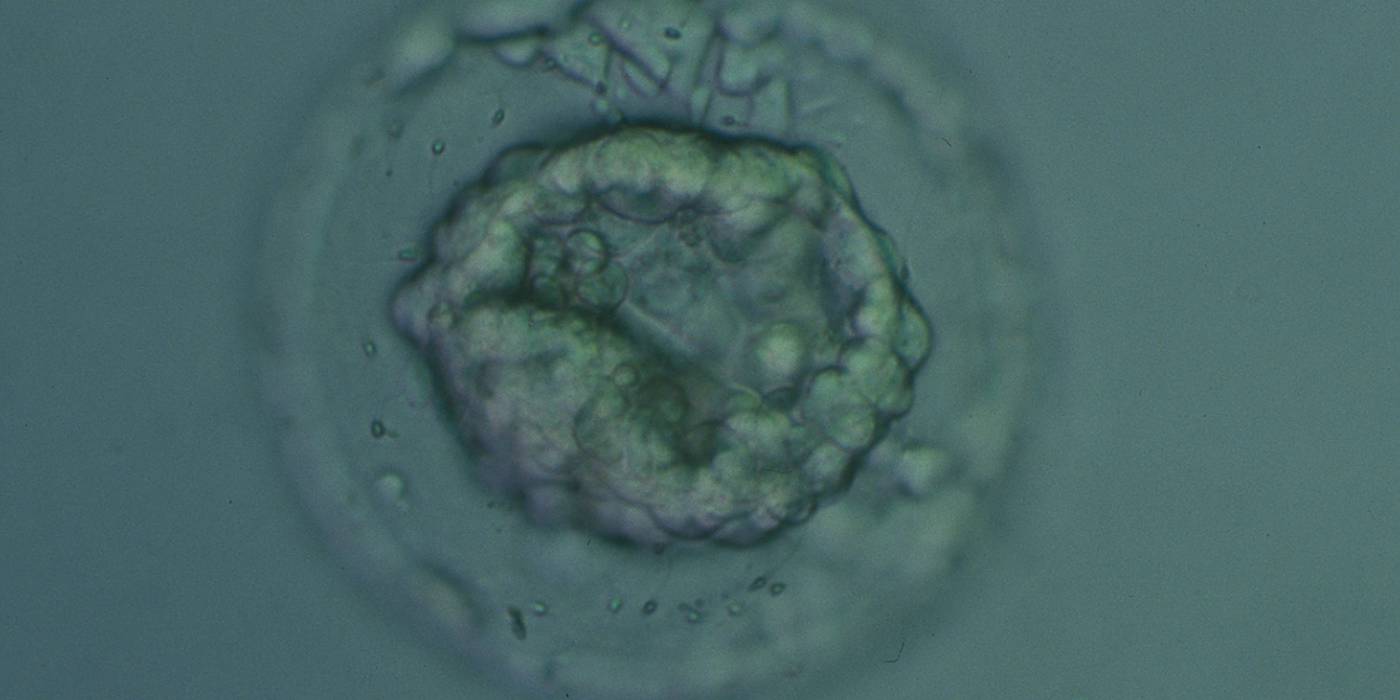
Figure 339
Collapsed blastocyst, which judging from the thickness of the ZP, is at least a Grade 3. The ICM and TE however cannot be accurately assessed. The blastocyst was transferred but the outcome is unknown.
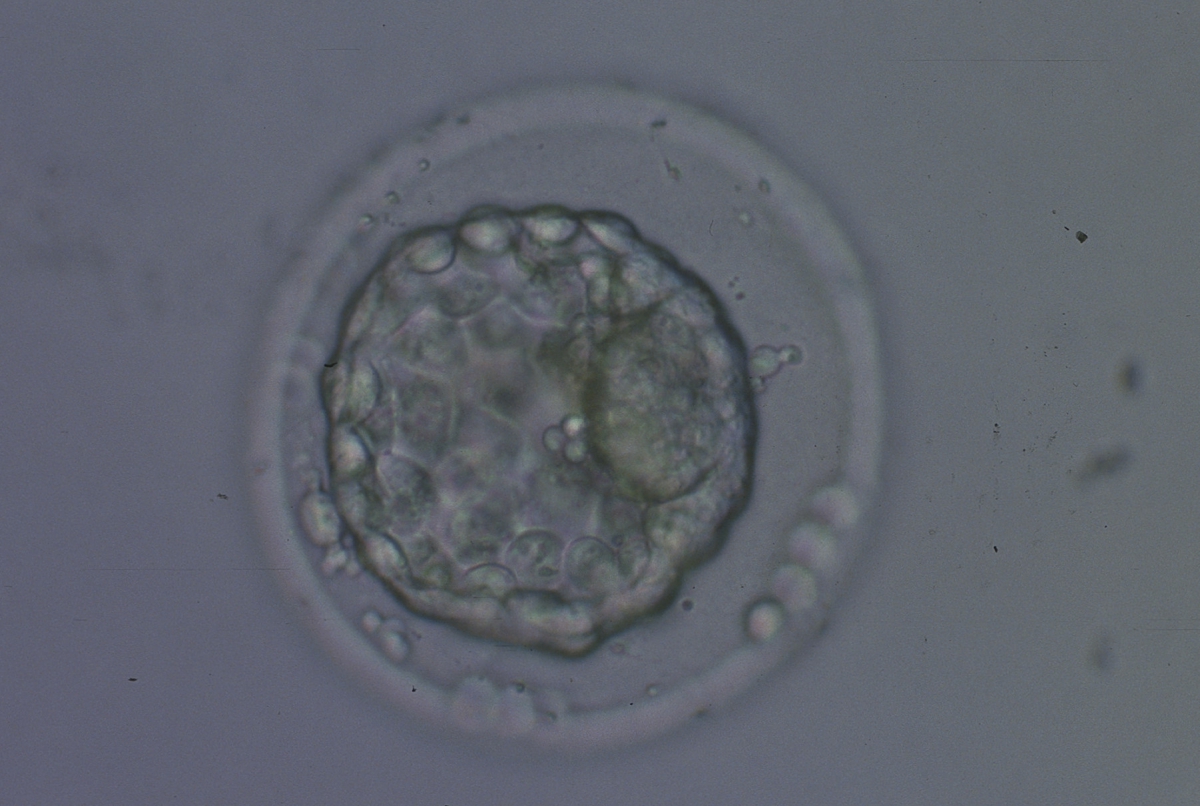
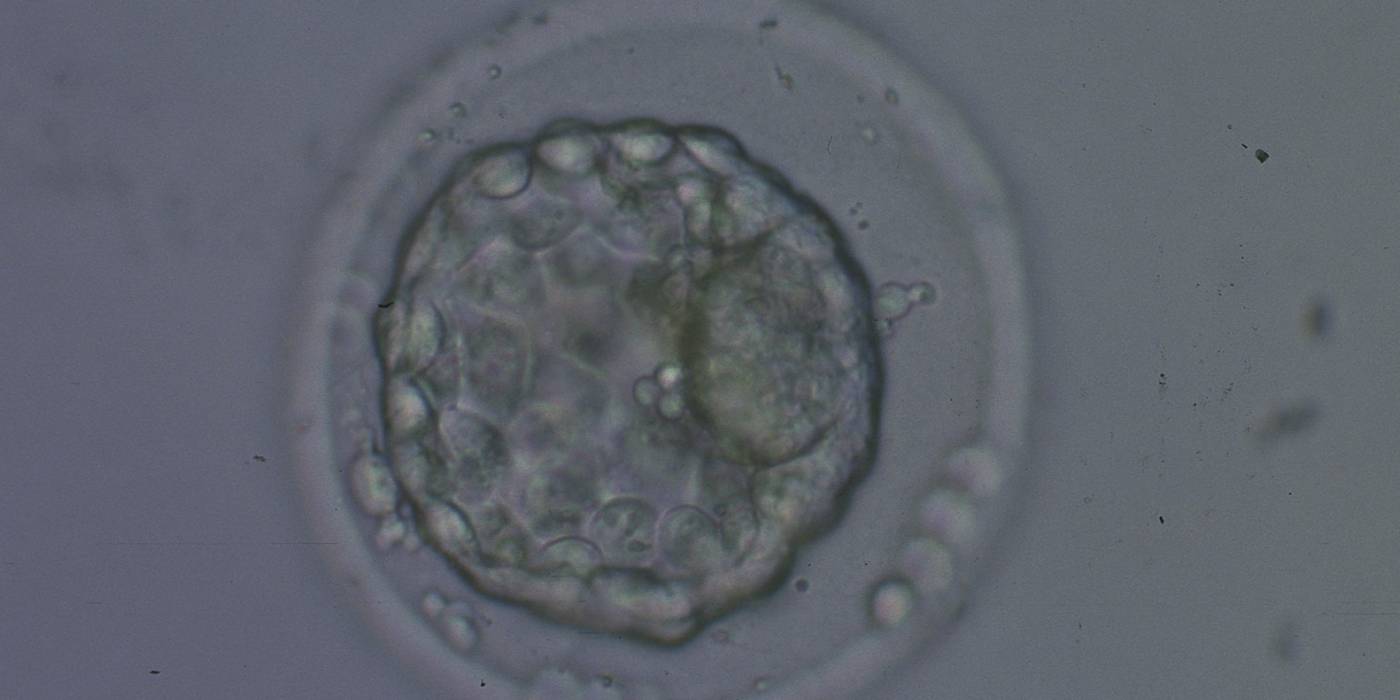
Figure 340
Collapsed blastocyst, which judging from the thickness of the ZP, is at least a Grade 3. In this view the ICM is clearly discernible at the 3 o'clock position and appears to be of Grade 1 quality. Similarly what can be seen of the TE cells appears to be Grade 1 in quality. There is some extraembryonic cellular debris in the PVS. The blastocyst was transferred but failed to implant.
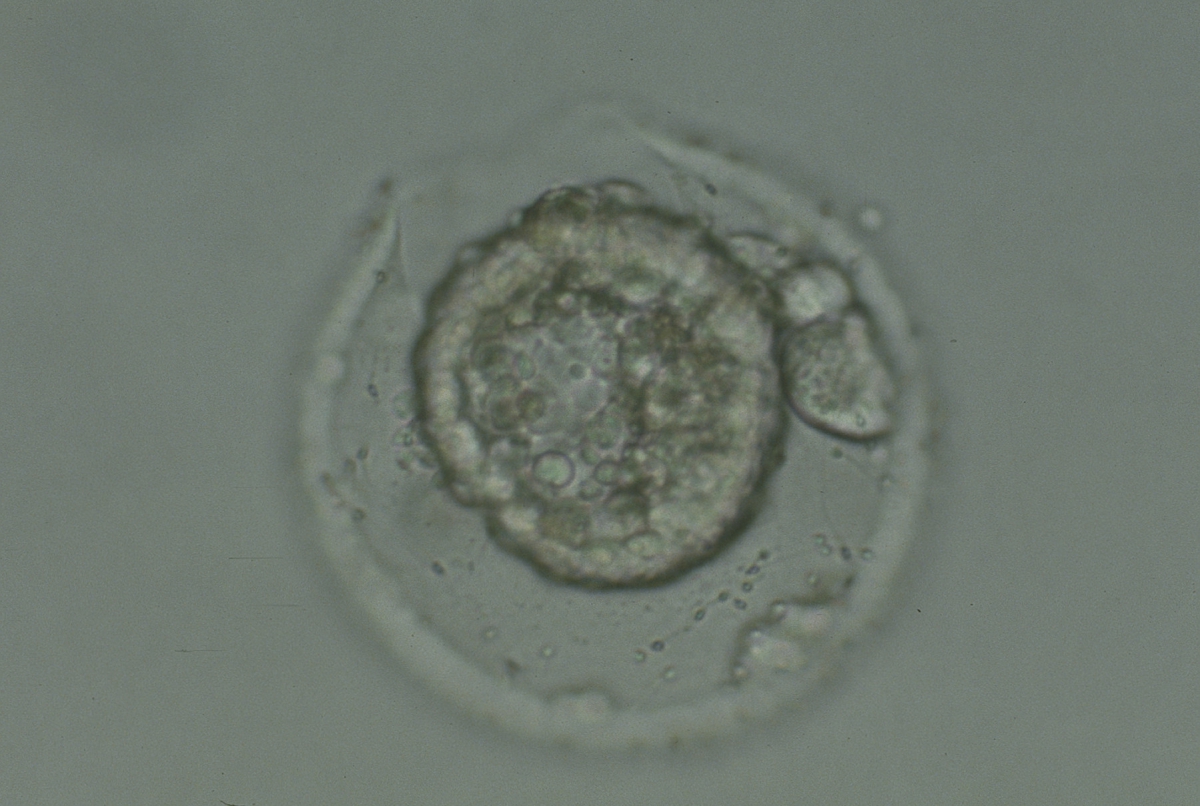
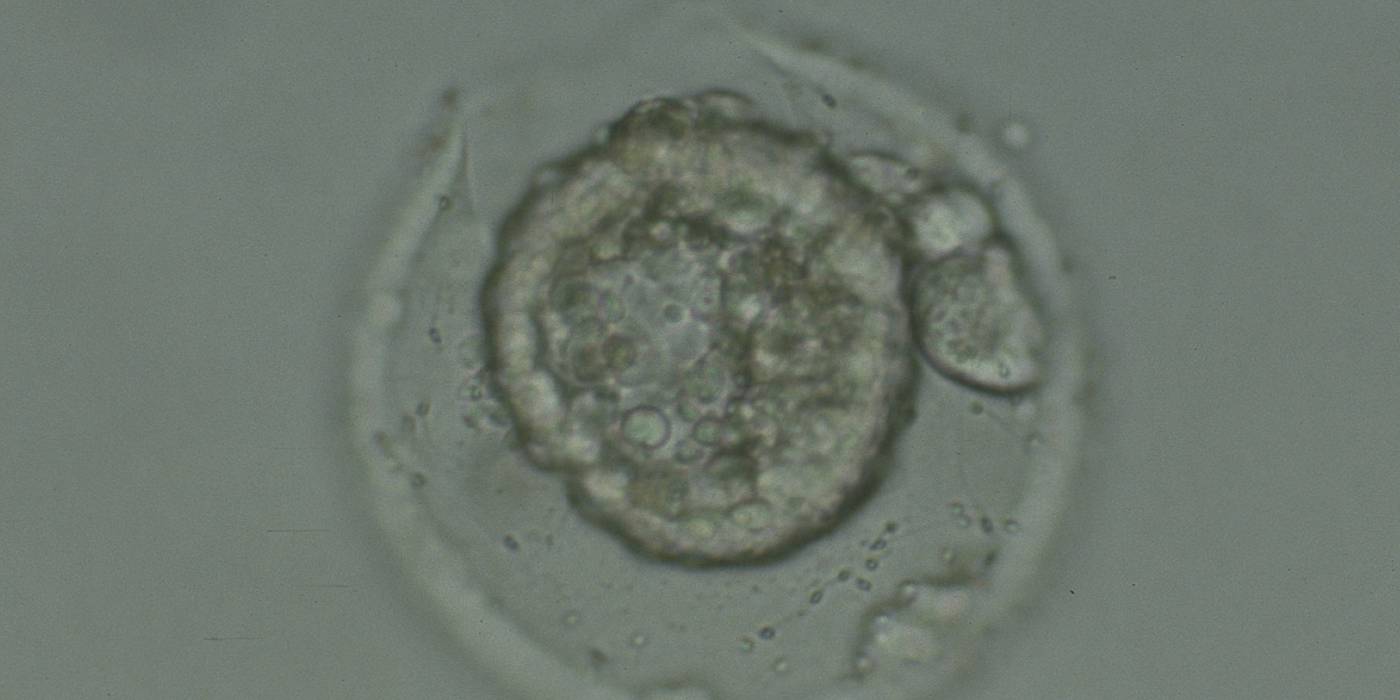
Figure 341
Collapsed hatching blastocyst (Grade 5) with the hatching site clearly visible as a breach in the ZP at the 11 o'clock position in this view. There is extraembryonic cellular debris both within the blastocoel cavity and external to the blastocyst in the PVS. Both the ICM and TE cannot be properly evaluated. The blastocyst was transferred but failed to implant.
The timing and grade of blastocyst expansion has been shown by several investigators to be an important predictor of implantation (Dokras et al., 1993; Gardner et al., 2000; Shapiro et al., 2008; Ahlström et al., 2011). It must, however, be remembered that embryologists have traditionally chosen to transfer the most developed blastocyst with the highest grade of expansion when available for transfer. No randomized, controlled trial comparing transfer of good quality blastocysts of a lower grade when they are present within a cohort that includes blastocysts with higher grades has been undertaken. Furthermore, recent evidence suggests that human female embryos undergo X chromosome inactivation from the 8-cell cleavage stage to the blastocyst stage of development which takes some time, therefore meaning that female blastocysts may be less expanded than their male sibling blastocysts but may be just as viable (van den Berg et al., 2009).
Article references:
Ahlström A, Westin C, Reismer E, Wikland M, Hardarson T. Trophectoderm morphology: an important parameter for predicting pregnancy and birth after single blastocyst transfer. Hum Reprod 2011;26:3289-3296.
Abstract/FREE Full Text
Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum Reprod 2011;26:1270-1283.
Abstract/FREE Full Text
Dokras A, Sargent IL, Barlow DH. Human blastocyst grading: an indicator of developmental potential? Hum Reprod 1993;8:2119-2127.
Abstract/FREE Full Text
Gardner DK, Schoolcraft WB. In vitro culture of human blastocysts. In: Jansen R, Mortimer D, editors. Toward Reproductive Certainty: Fertility and Genetics Beyond 1999. UK: Parthenon Publishing London; 1999. p. 378-388.
Google Scholar
Gardner DK, Lane M, Stevens J, Schlenker T, Schoolcraft WB. Blastocyst score affects implantation and pregnancy outcome: towards a single blastocyst transfer. Fertil Steril 2000;73:1155-1158.
CrossRef | Medline | Web of Science | Google Scholar
Hardarson T, Caisander C, Sjögren A, Hanson C, Hamberger L, Lundin K. A morphological and chromosomal study of blastocysts developing from morphologically suboptimal human preembryos compared to control blastocysts. Hum Reprod 2003;18:399-407.
Abstract/FREE Full Text
Shapiro BS, Daneshmand ST, Garner FC, Aguirre M, Thomas S. Large blastocyst diameter, early blastulation, and low preovulatory serum progesterone are dominant predictors of clinical pregnancy in fresh autologous cycles. Fertil Steril 2008;90:302-309.
CrossRef | Medline | Web of Science | Google Scholar
Van den Berg IM, Laven JS, Stevens M, Jonkers I, Galjaard RJ, Gribnau J, van Doorninck JH. X chromosome inactivation is initiated in human preimplantation embryos. Am J Hum Genet 2009;84:771-779.
CrossRef | Medline | Web of Science | Google Scholar
Watson AJ, Natale DR, Barcroft LC. Molecular regulation of blastocyst formation. Anim Reprod Sci 2004;82–83:583-592.
Google Scholar
