C. Pronuclear morphology
C.1 Alignment parallel/tangential to the plane of the polar bodies
With the extrusion of the second polar body, following sperm entry and oocyte activation, the polar axis is established. Correct alignment of PNs onto this axis is necessary for the formation of polar axes at syngamy and subsequent completion of the first cleavage division and normal development (Edwards and Beard, 1997; Payne et al., 1997; Garello et al., 1999; Gardner, 2001; Scott, 2001). At apposition, the chromatin of both PNs begins to polarize and rotate to face each other with the male PN rotating onto the female PN and placing the centrosome into the furrow between the two PNs (Van Blerkom et al., 1997). In this way, the longitudinal axis of PNs is parallel to the plane of polar bodies (Figs 111–115). Further rotation brings the PNs aligned onto the polar axis; the position of the second polar body defines the plane of the first cleavage division (Figs 116–121). All these movements and rotations could cause the formation of the clear cortical zone known as the halo (Figs 114, 118 and 119).
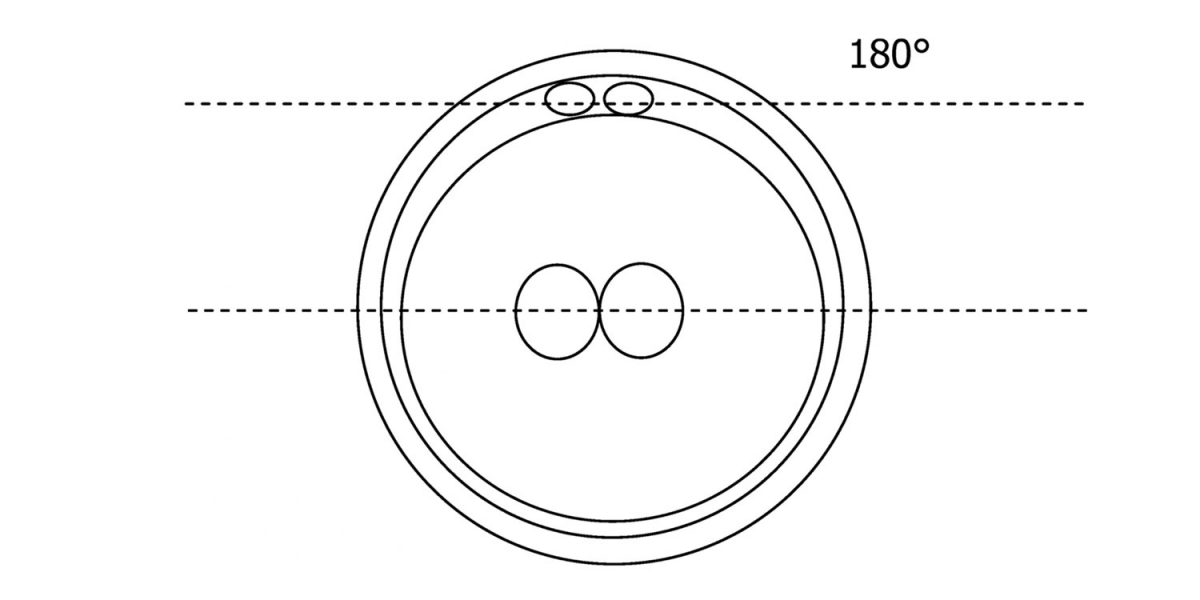
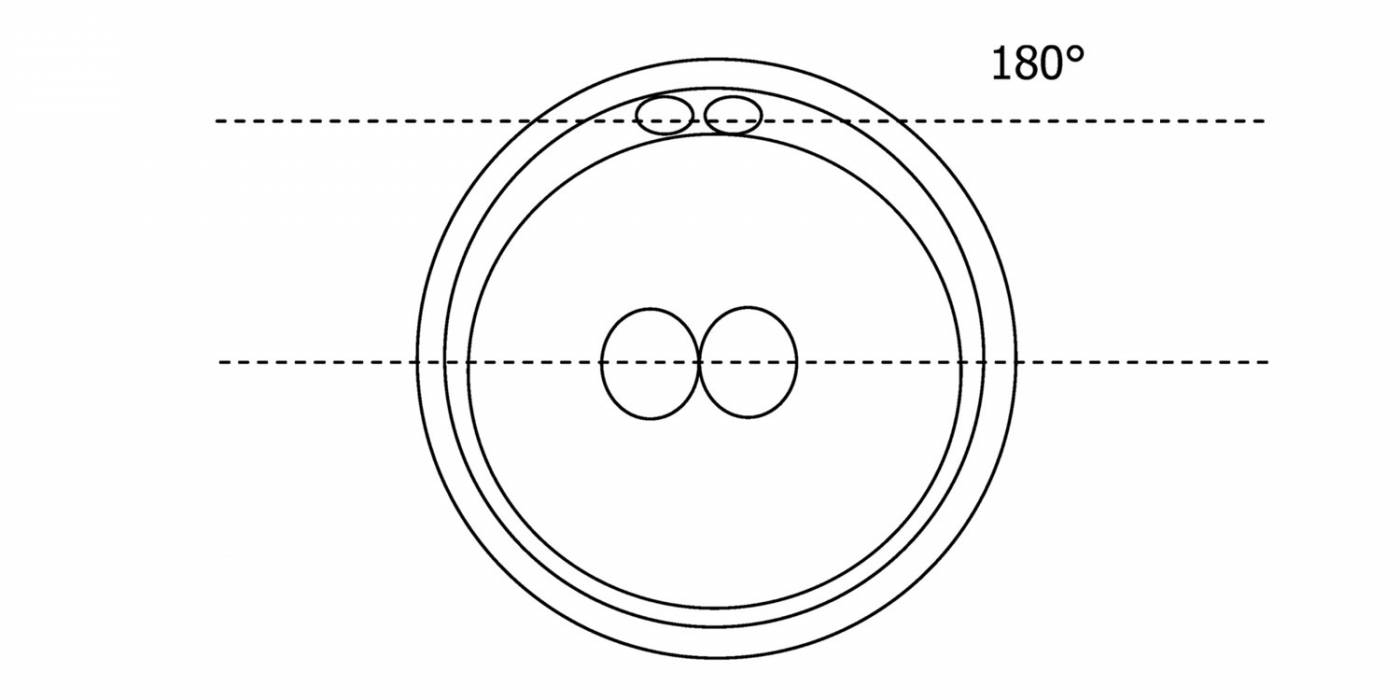
Figure 111
Diagram showing the plane through the polar bodies which is parallel to the contrasting plane through the longitudinal axis of the PNs.
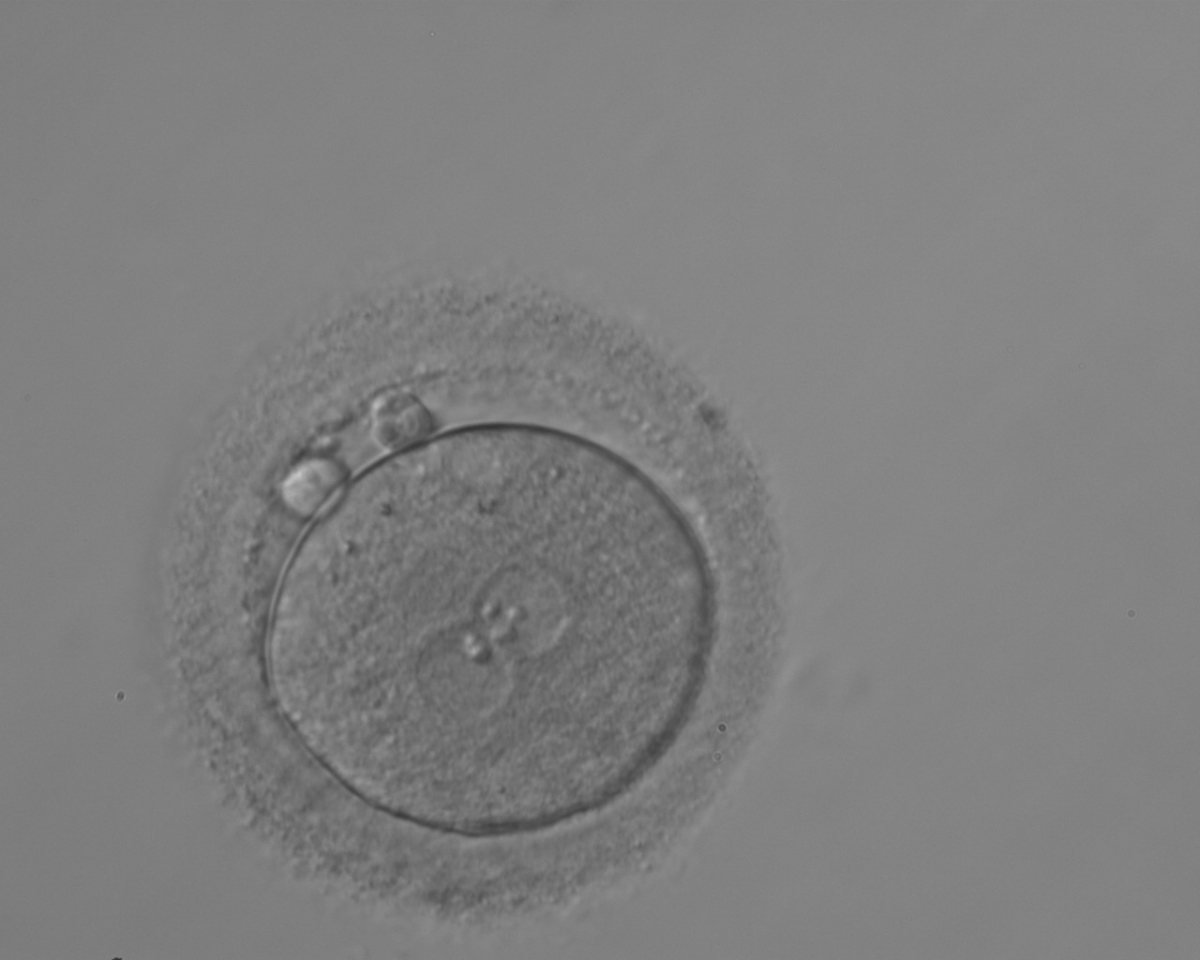
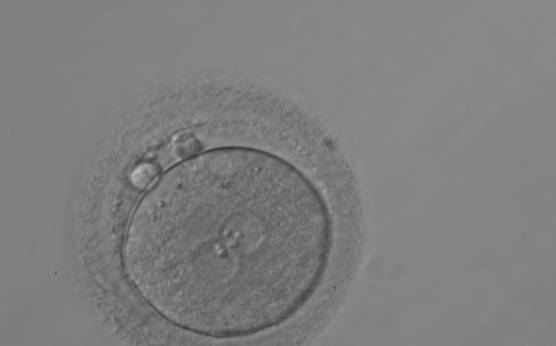
Figure 112
A zygote generated by ICSI with different number of large-sized NPBs aligned at the PN junction (200× magnification). PNs are juxtaposed and centralized; polar bodies are located parallel to the longitudinal axis through the PNs.
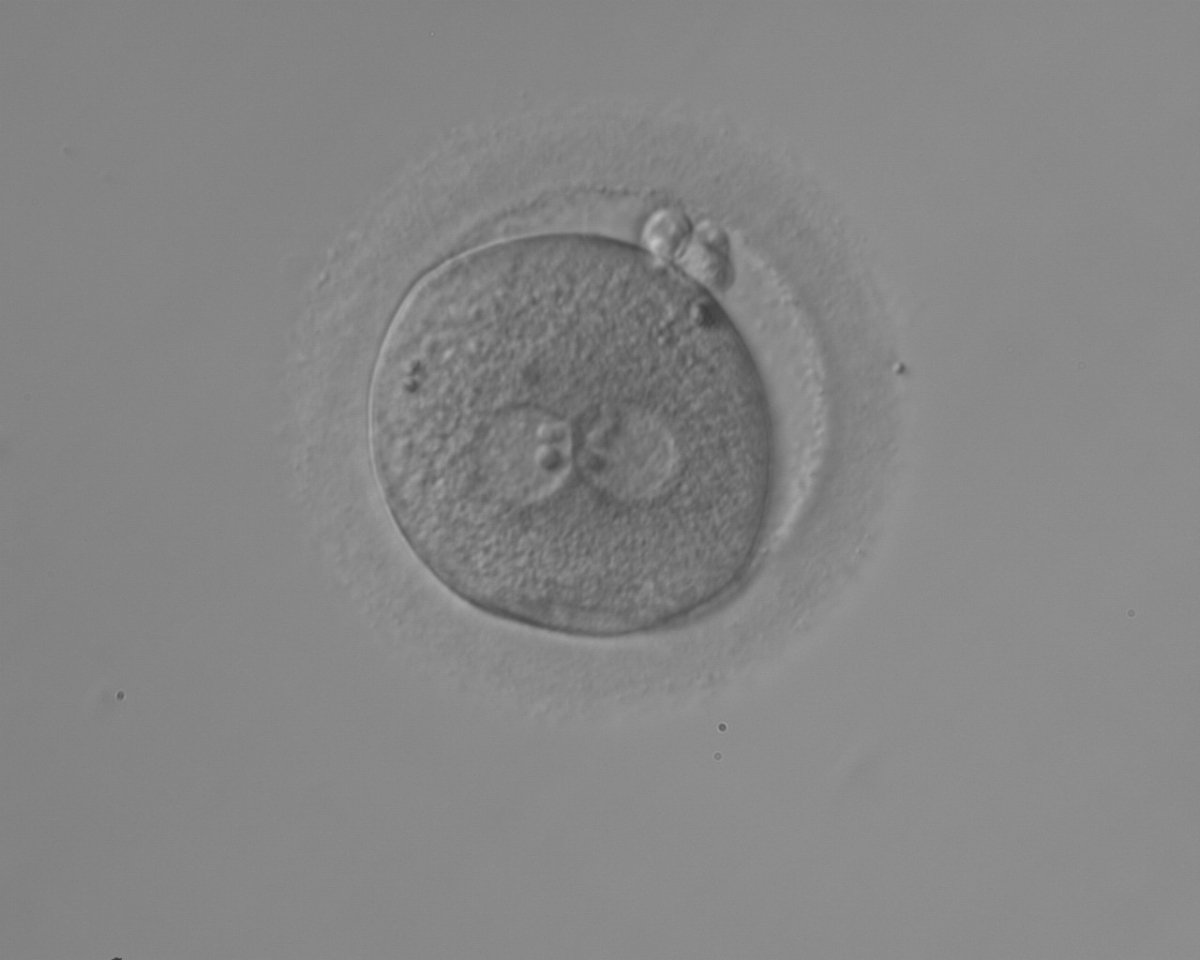
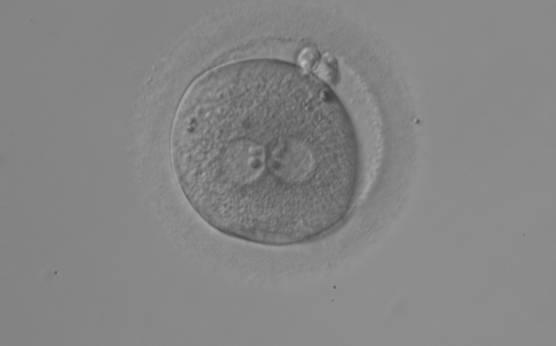
Figure 113
An ICSI zygote with large-sized NPBs aligned at the PN junction (200× magnification). PNs are juxtaposed and centralized, fragmented polar bodies are located parallel to the longitudinal axis through the PNs (±30°).
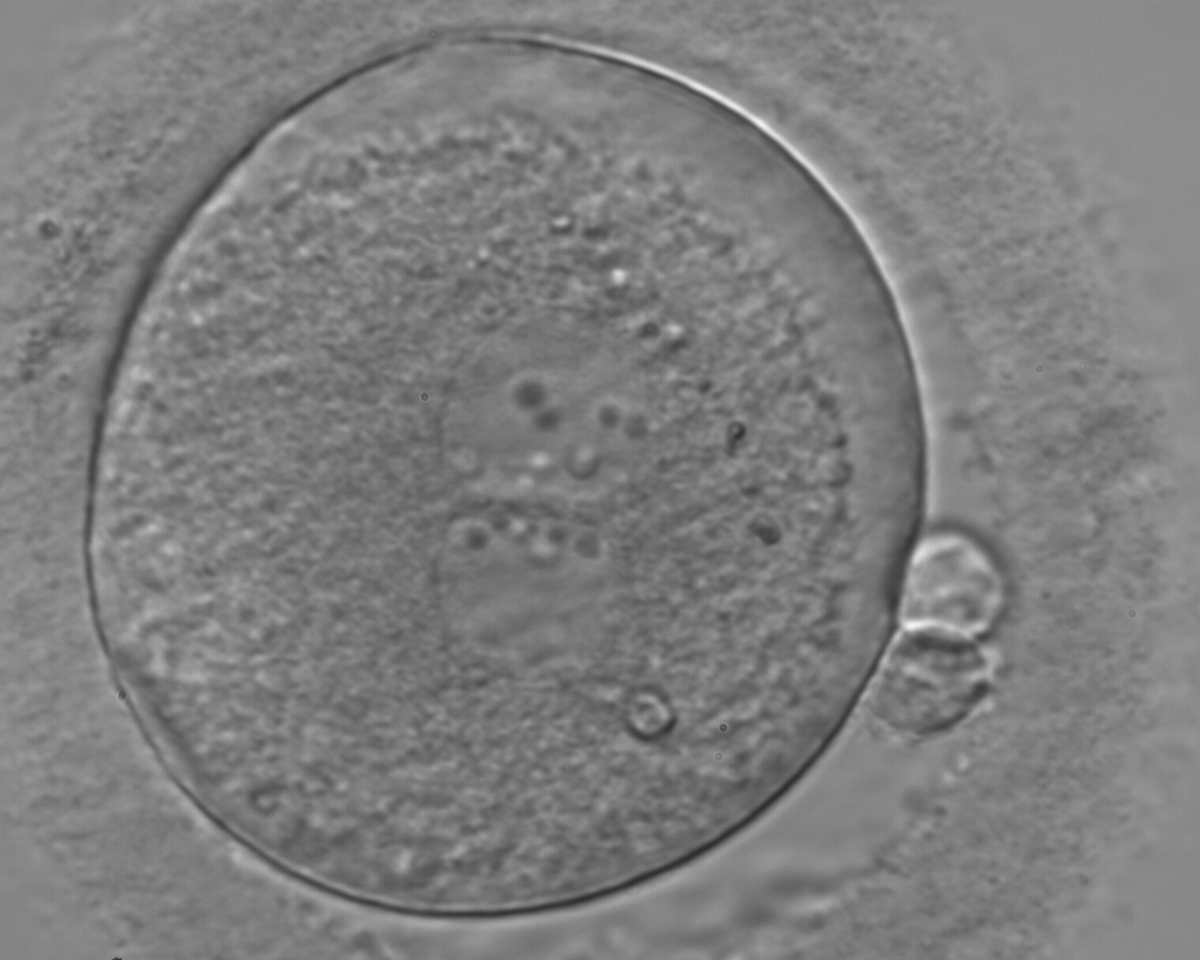
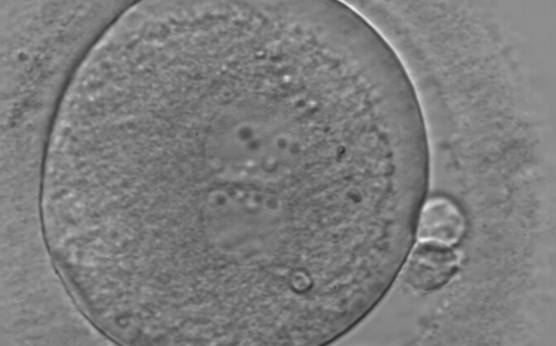
Figure 114
An ICSI zygote displaying juxtaposed and centralized PNs. PN alignment is parallel to the plane of the polar bodies. The zygote displays inequality in number and alignment of NPBs. The latter are aligned in one PN and scattered in the other one with respect to the PN junction. A clear cortical zone with some inclusion bodies immediately below the area can be noted. It was replaced on Day 3 (eight cell cleavage embryo) but failed to implant.
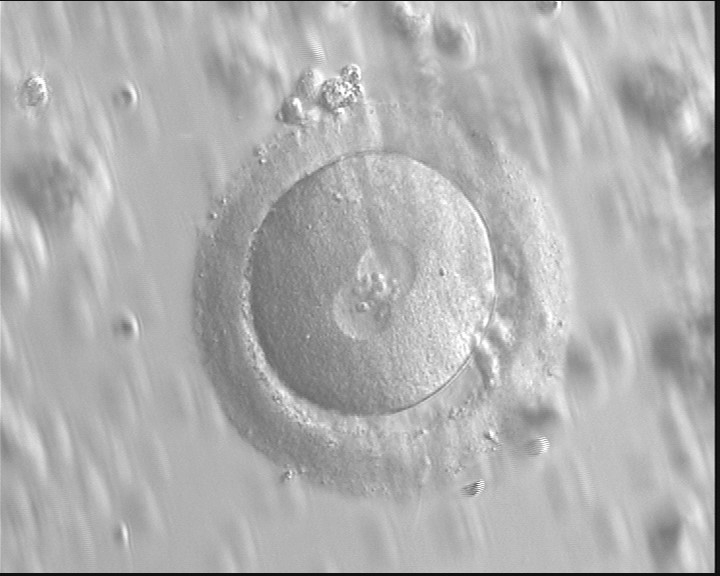
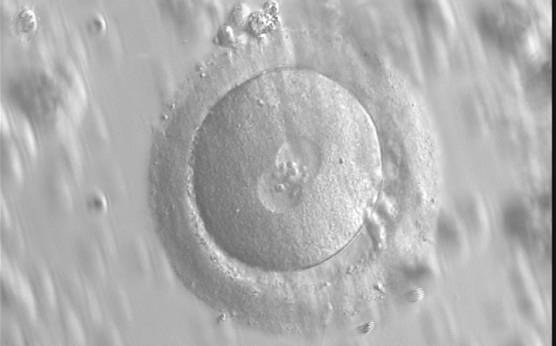
Figure 115
A zygote with an equal number of large-sized NPBs aligned at the PN junction (400× magnification). PNs are juxtaposed and centralized and the polar bodies are located parallel to the longitudinal axis through the PNs. Observed 16 h after IVF.
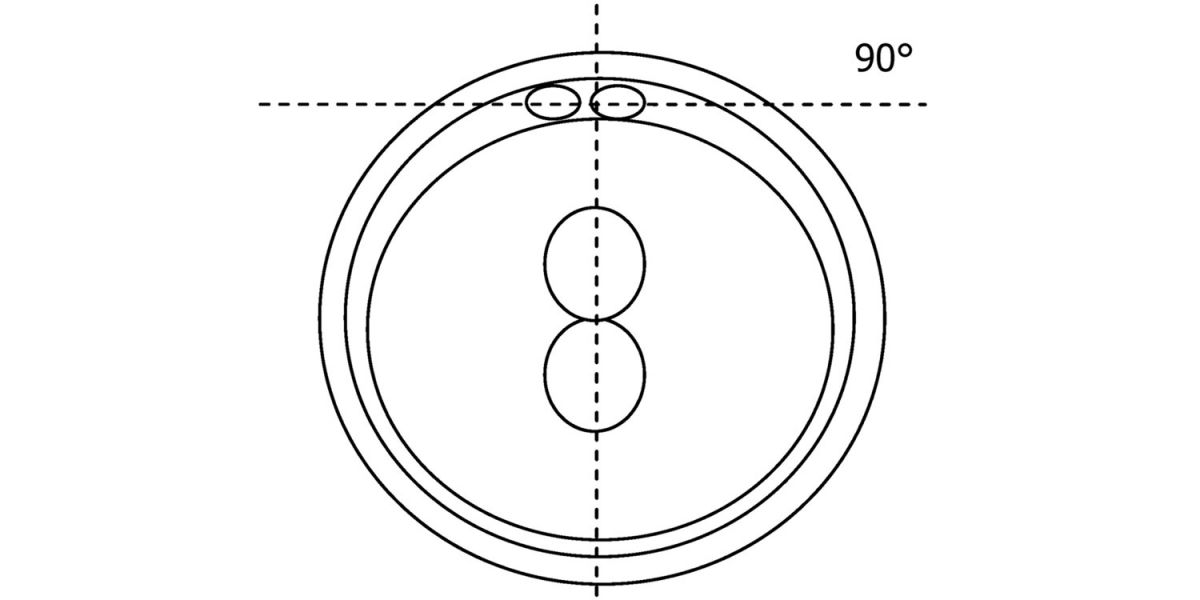
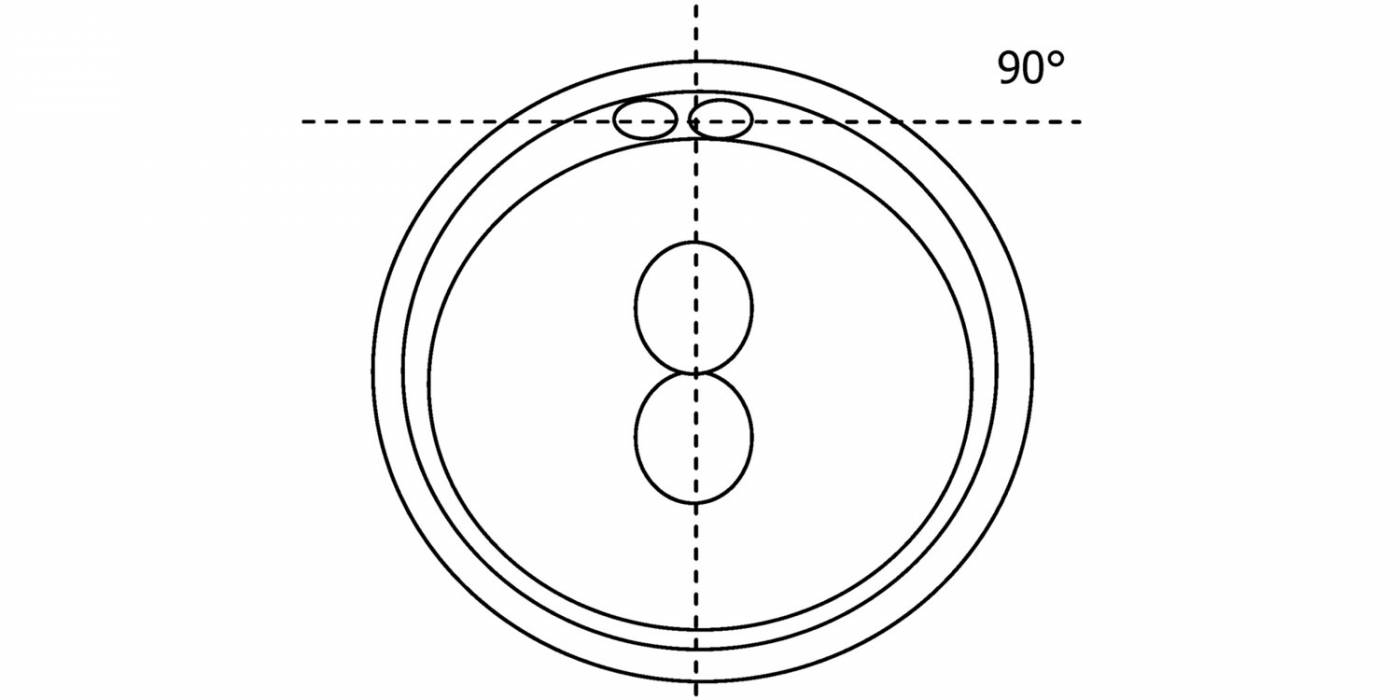
Figure 116
Diagram showing the plane through the polar bodies which is tangential to the contrasting plane through the longitudinal axis of the PNs.
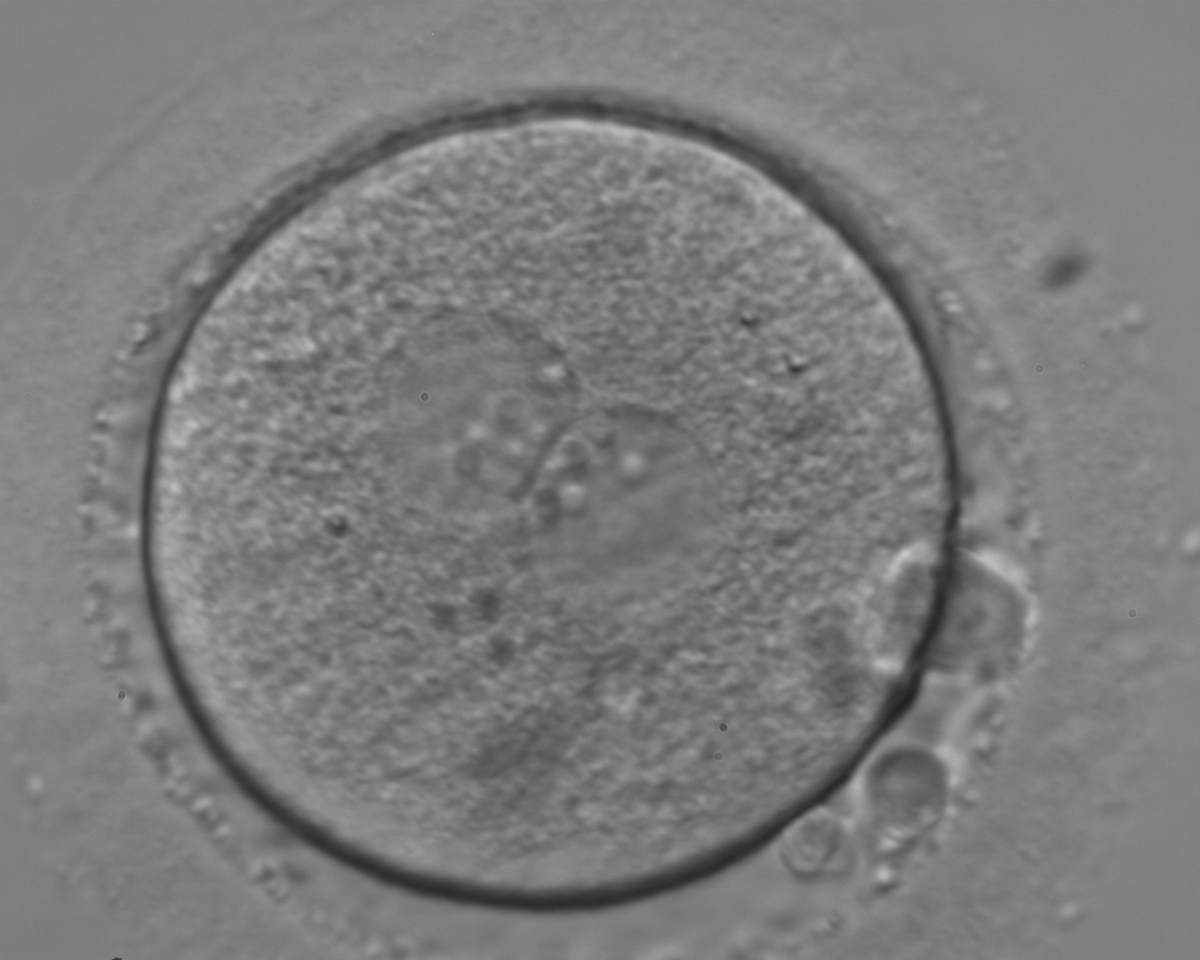
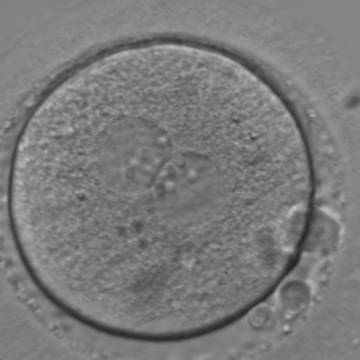
Figure 117
An ICSI zygote with large-sized NPBs aligned at the PN junction (400× magnification). PNs are juxtaposed and centralized; polar bodies (the first polar body is fragmented, the second is intact) are located tangential to the longitudinal axis through the PNs.
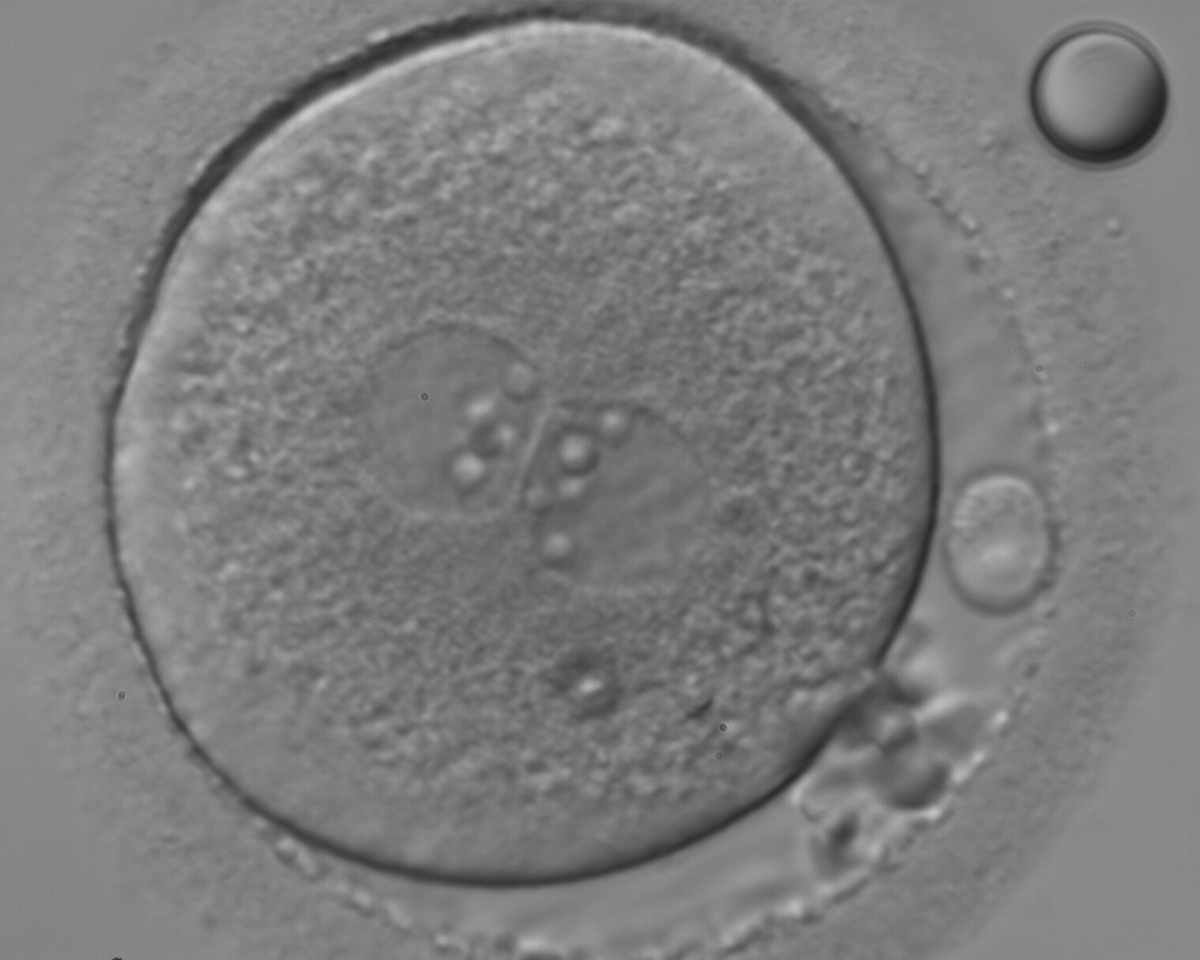
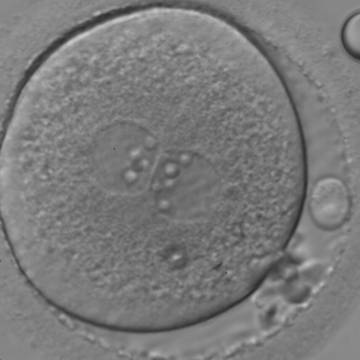
Figure 118
An ICSI zygote with large-sized NPBs aligned at the PN junction (400× magnification). PNs are juxtaposed and centralized; polar bodies (the first polar body is fragmented, while the second is intact) are located tangential to the longitudinal axis through the PNs.
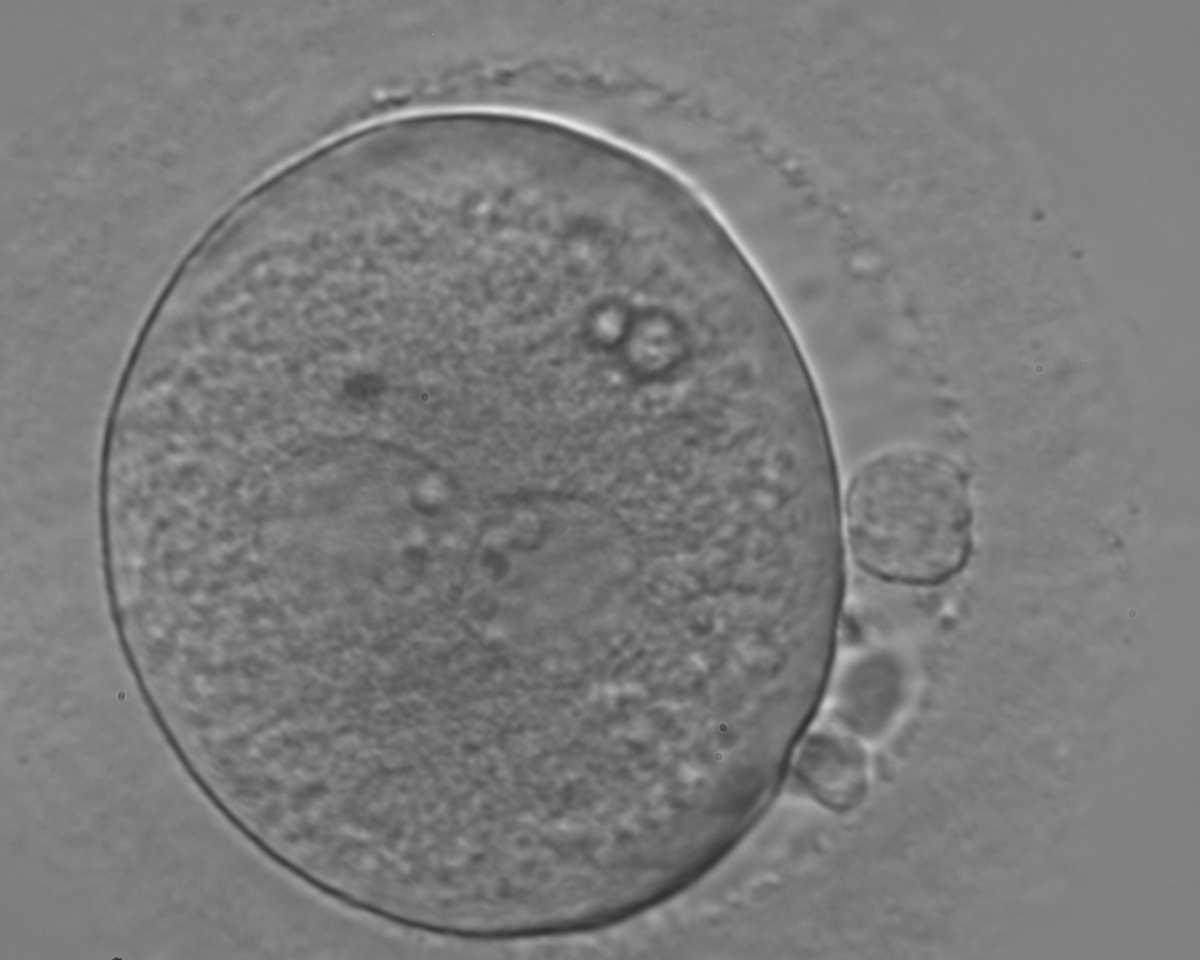
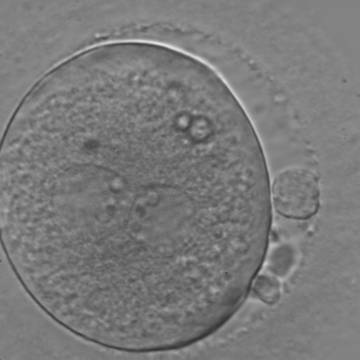
Figure 119
An ICSI zygote with large-sized NPBs aligned at the PN junction (200× magnification). PNs, juxtaposed and centralized, are tangential to the plane through the polar bodies. It was transferred on Day 3 (seven cells) along with two other embryos but failed to implant.
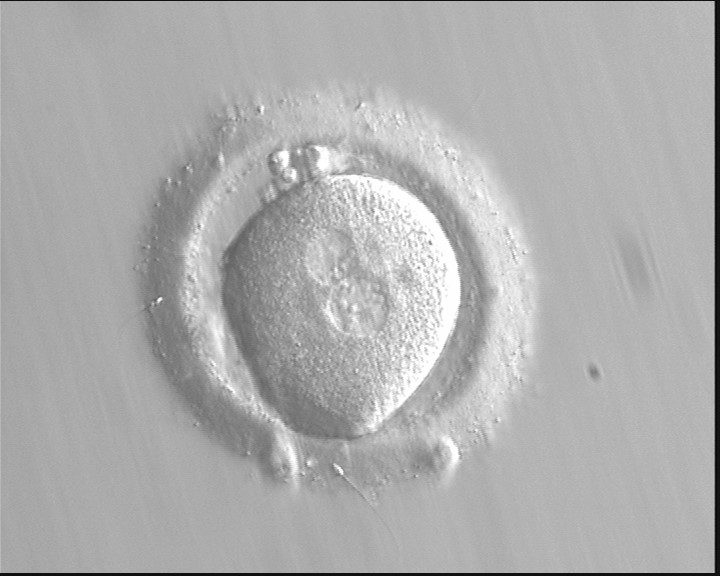
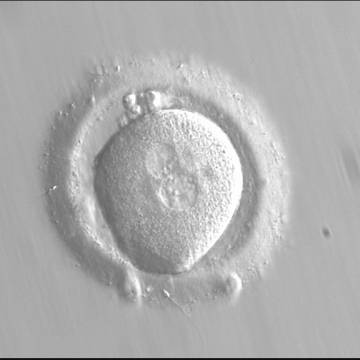
Figure 120
Zygote observed at 16.5 h post-IVF displaying unequal-sized juxtaposed and centralized PNs aligned tangentially to the plane through the polar bodies (400× magnification). NPBs are scattered in one PN and aligned in the other. The oocyte is irregular in shape and the PVS is enlarged. It failed to implant following transfer.
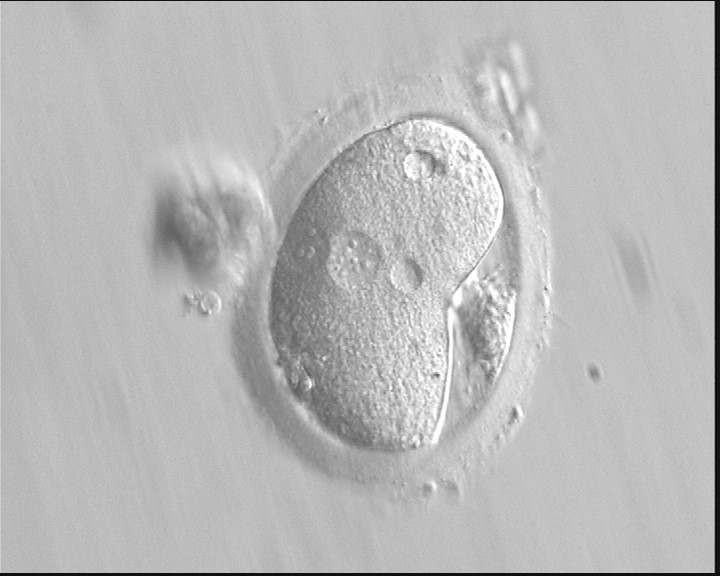
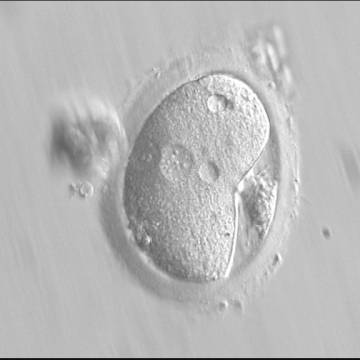
Figure 121
A deformed zygote with peripheral PNs and polar bodies aligned tangentially to the plane through the polar bodies observed 15 h post-IVF (400× magnification). Polar bodies are highly dysmorphic. Note the presence of small vacuoles in the cytoplasm.
C.2 Large angle between polar bodies
The failure of PNs to be juxtaposed and centrally positioned within the cytoplasm or having non-aligned polar bodies with respect to PNs at fertilization check could result in altered development, i.e. fertilization failure and abnormal cleavage of the embryo (Gianaroli et al., 2003; Alpha Scientists in Reproductive medicine and ESHRE Special Interest Group of Embryology, 2011). Large angles between polar bodies (Figs 122–124) have been suggested to be predictors of poor embryo development (Gianaroli et al., 2003). This effect could be due to sub-optimal orientation of PNs (Figs 125 and 126) generating a great degree of cytoplasmic turbulence, which could facilitate uneven cleavage or fragmentation (Garello et al., 1999).
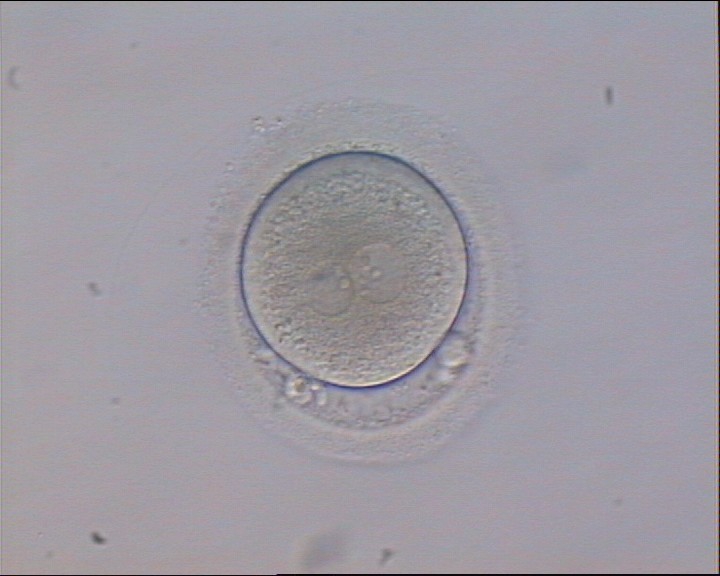
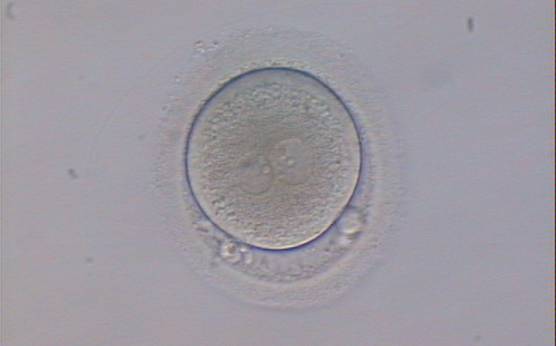
Figure 122
An ICSI zygote displaying two PNs of approximately equal size, juxtaposed and clearly visible in the middle of the cytoplasm (400× magnification). Polar bodies are rotated >30° off the longitudinal axis through the PNs. A clear cortical area and coarse granularity of the cytoplasm can be observed. It was transferred but failed to implant.
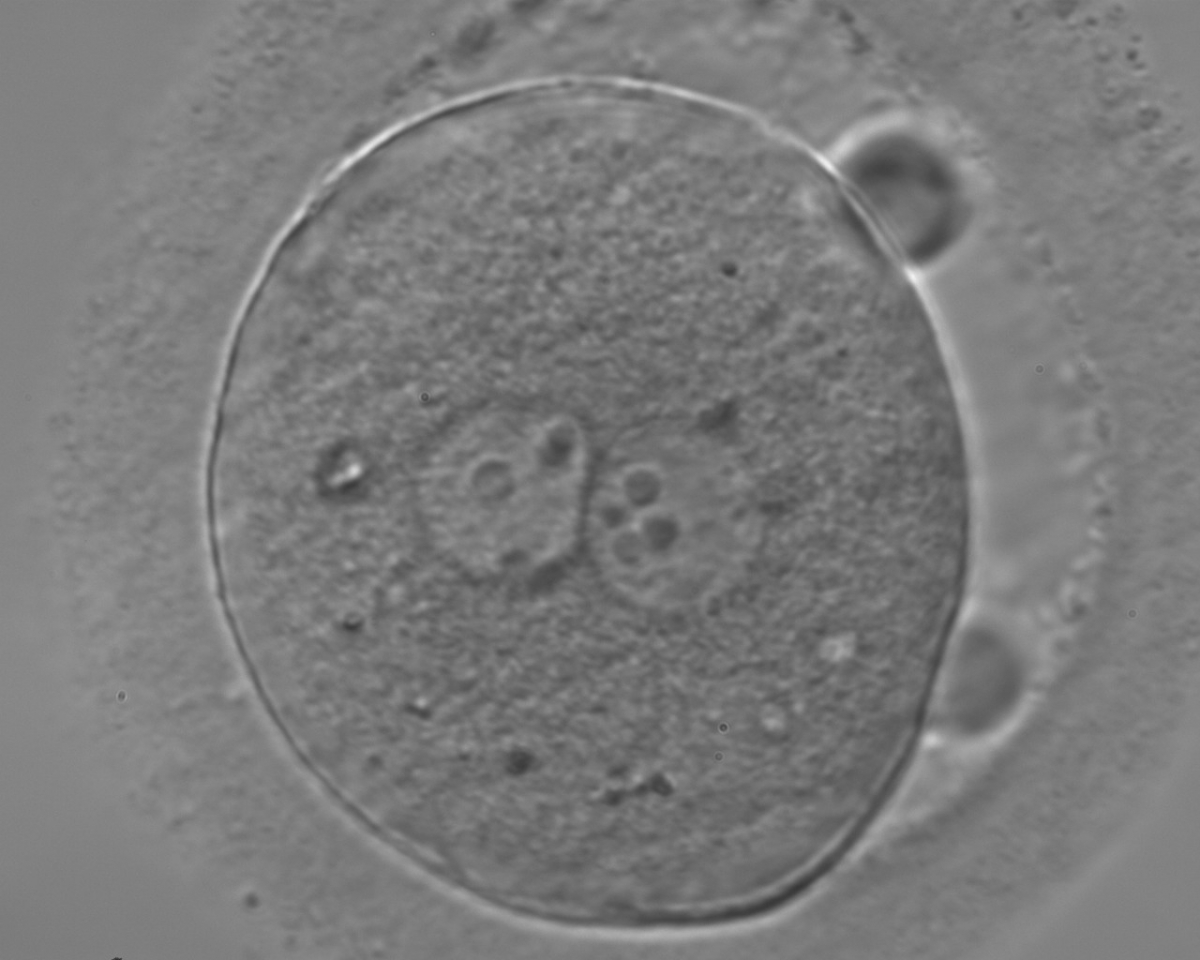
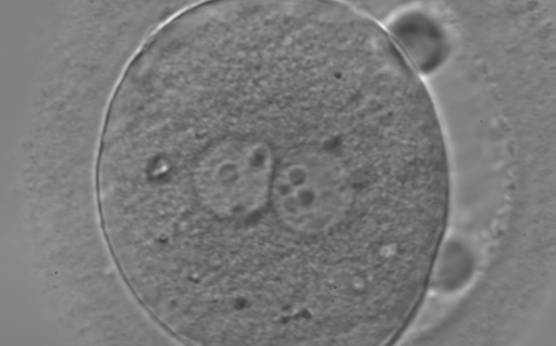
Figure 123
Zygote observed 18 h post-ICSI, with inequality in number and alignment of the NPBs (400× magnification). PNs are juxtaposed and centralized, NPBs are aligned in one of the two PNs and scattered in the other with respect to the PN junction. Both polar bodies are rotated >30° off the longitudinal or meridional axis with a large degree of separation between them. It is possible to observe a clear cortical zone and some dark inclusion bodies. It was transferred and failed to implant.
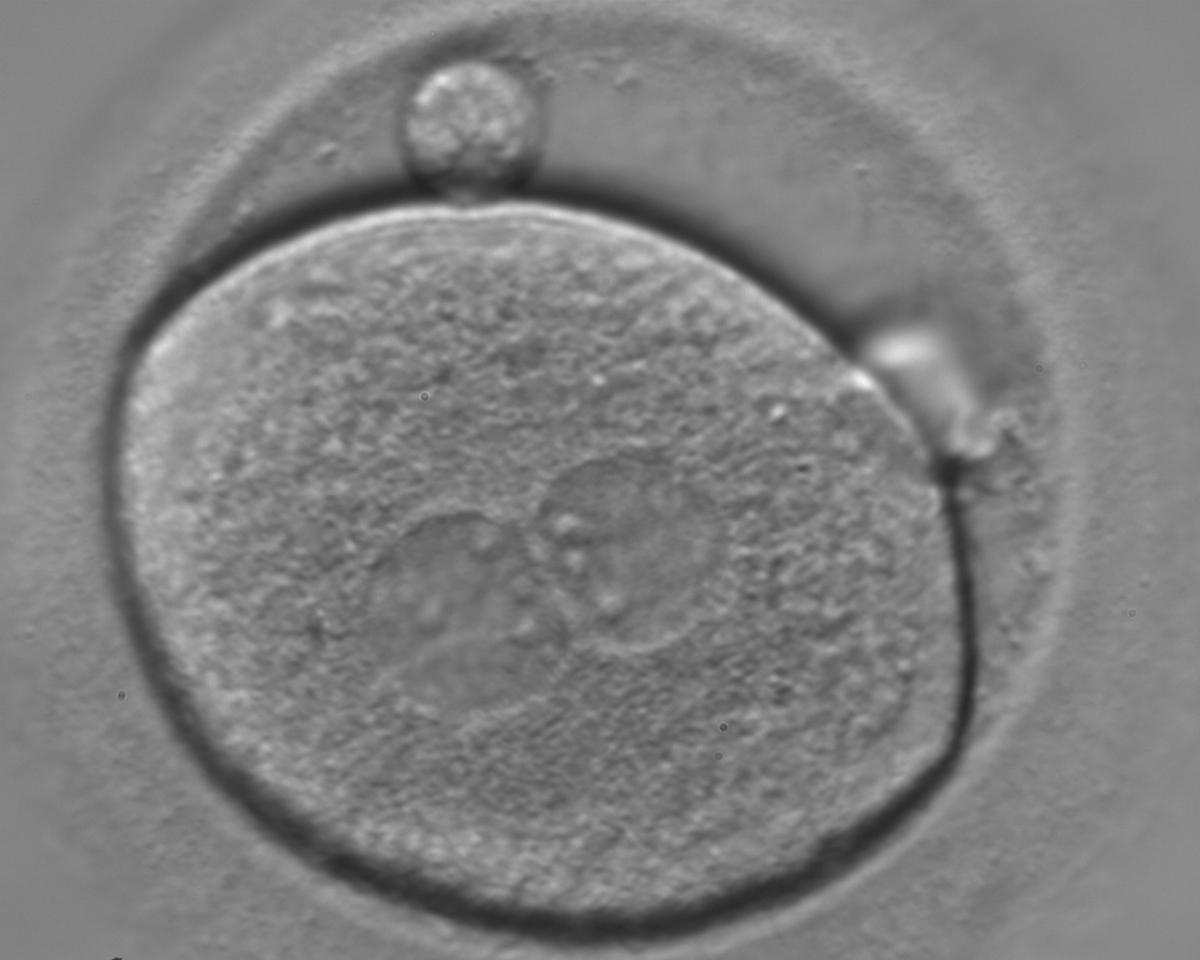
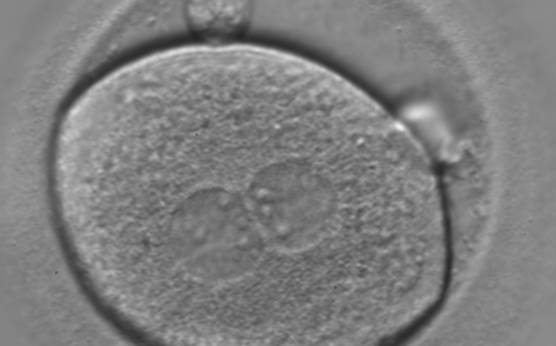
Figure 124
A zygote of irregular shape observed 18 h post-ICSI (400× magnification). The visible PNs show unequal distribution of the NPBs. PNs are juxtaposed and centralized and the polar bodies form a great angle with respect to the longitudinal axis through the PNs. It was transferred but failed to implant.
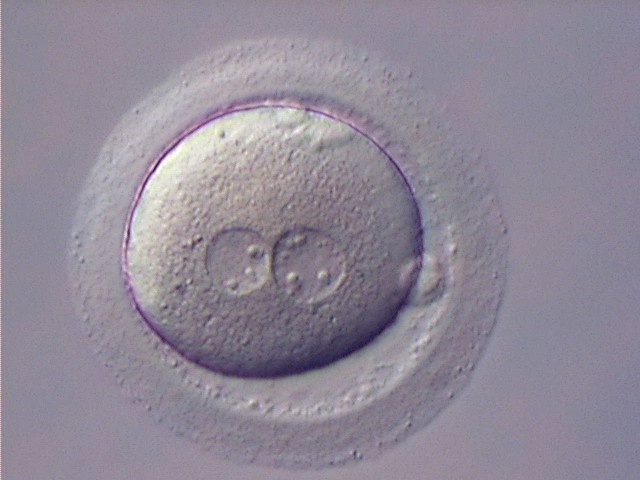
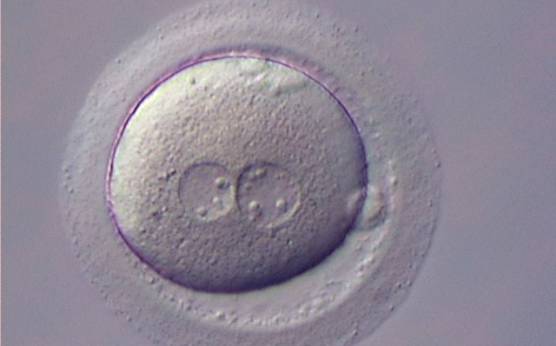
Figure 125
Zygote with large-sized NPBs scattered with respect to the PN junction (400× magnification). A clear cortical zone can be observed in the cytoplasm and the ZP is thick and very dense. Polar bodies form a great angle with respect to the longitudinal axis through the PNs. It was transferred but implantation outcome is unknown.
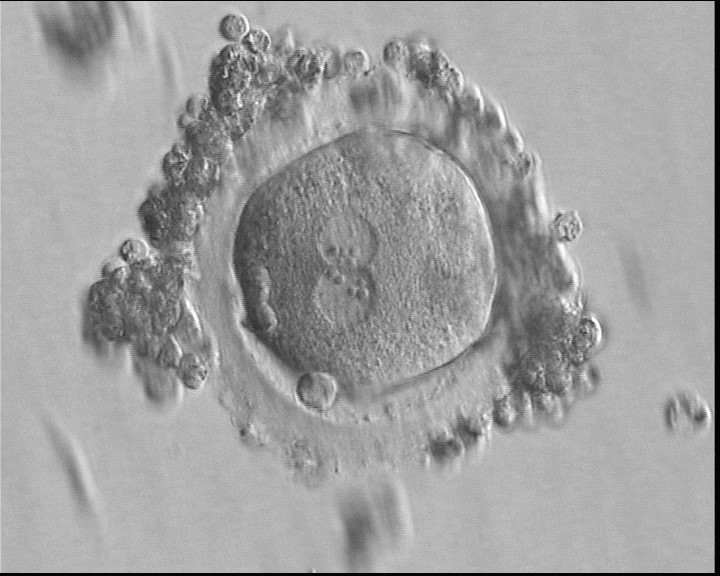
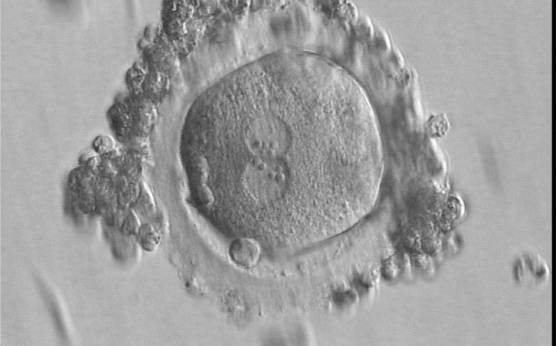
Figure 126
A zygote of irregular shape 16.5 h after IVF in which two PNs are clearly visible with NPBs aligned in both (400× magnification). Polar bodies form a right angle: one is aligned with the longitudinal axis through the PNs and the other is aligned with the meridional axis. It was transferred but failed to implant.
C.3 Abutment/separation between PNs
In human oocytes, the aster from the sperm centrosome organizes the microtubules, which control the abutment and apposition of PNs (Figs 127–131), and direct the formation of polar axes at syngamy by setting the plane of the first division. The subsequent movements and rotations favour the distribution of the mitochondria and chromatin alignment, which are essential for correct development.
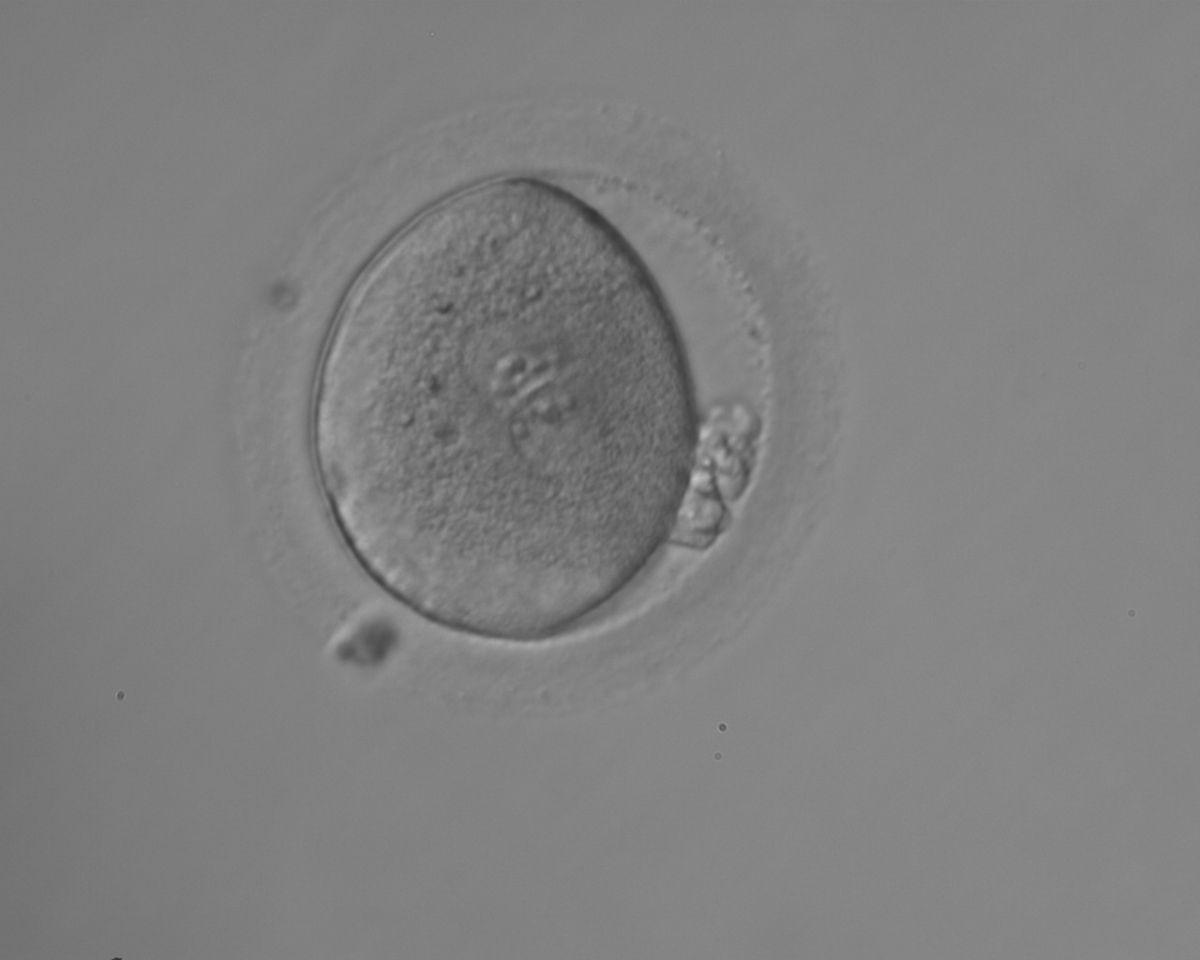
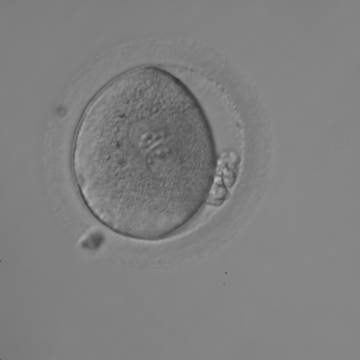
Figure 127
Irregularly shaped zygote showing two centrally located and juxtaposed PNs with equal numbers of large-sized NPBs aligned at the PN junction (200× magnification). Fragmented polar bodies are located parallel to the longitudinal axis through the PNs (±30°). A clear cortical area and coarse granularity of the cytoplasm can be observed. It was transferred on Day 3 along with two other embryos and the patient delivered two healthy baby girls and one healthy baby boy.
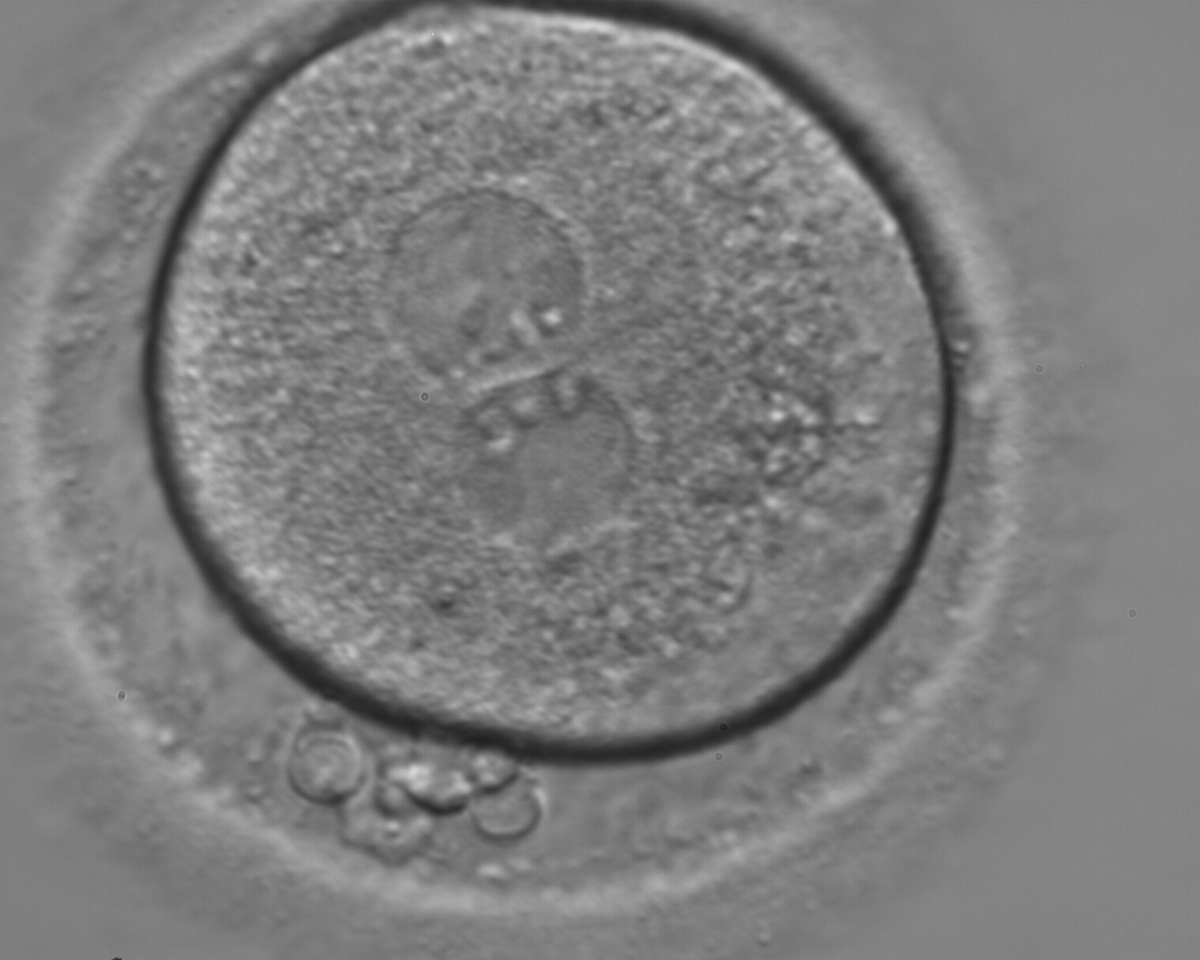
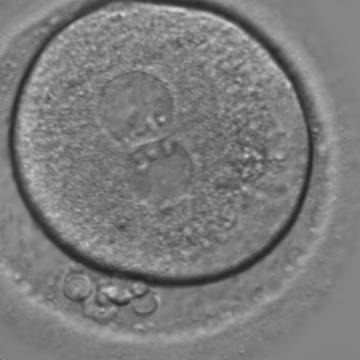
Figure 128
A zygote with two centrally located and juxtaposed PNs, an enlarged PVS and a significant amount of debris in the PVS (400× magnification). It was transferred but the result is unknown.
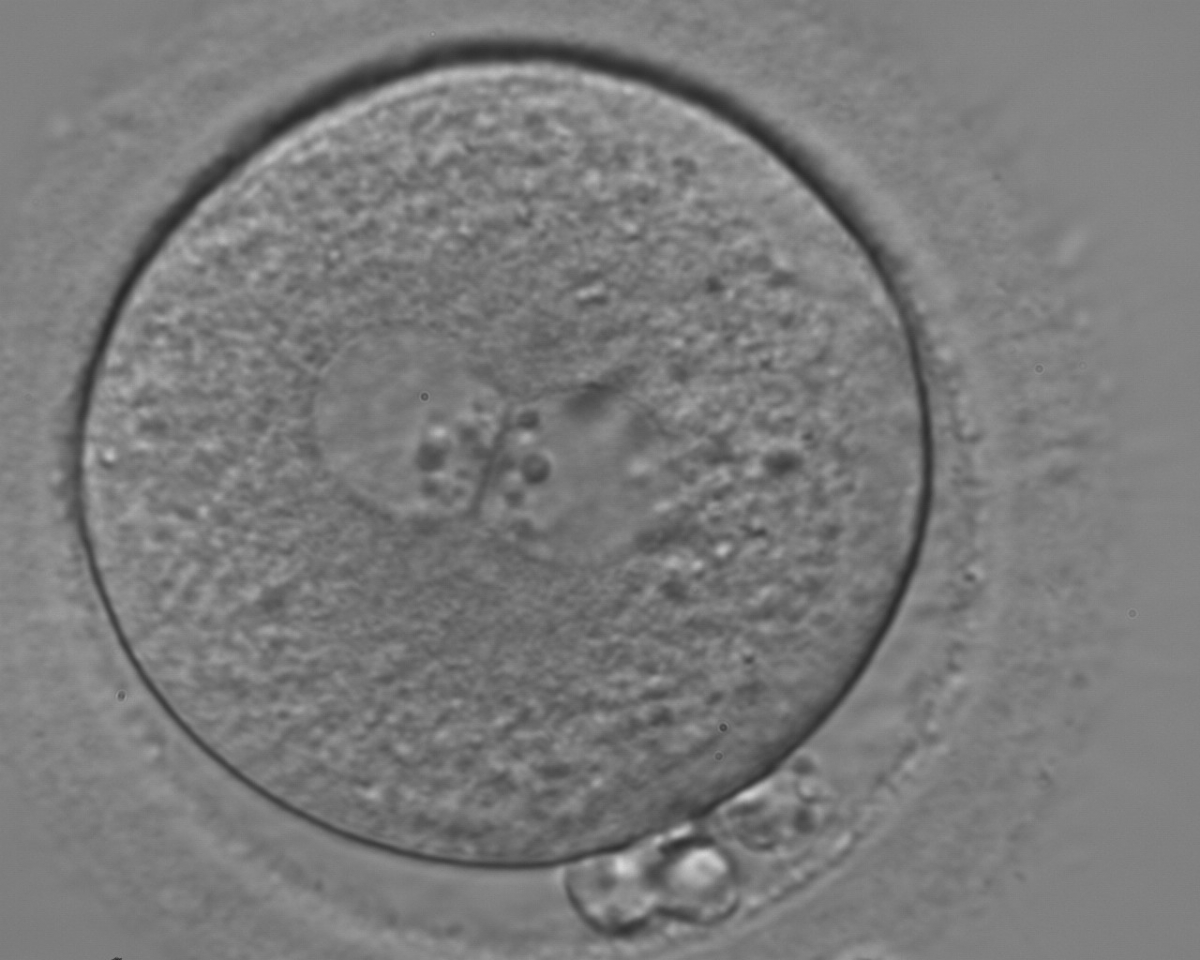
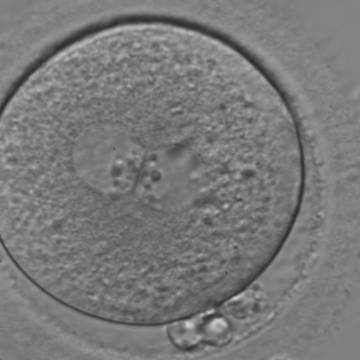
Figure 129
A zygote with two centrally located and perfectly juxtaposed PNs in a granular cytoplasm with a clear cortical zone (400× magnification). NPBs are aligned, but are different in size. It was transferred but failed to implant.
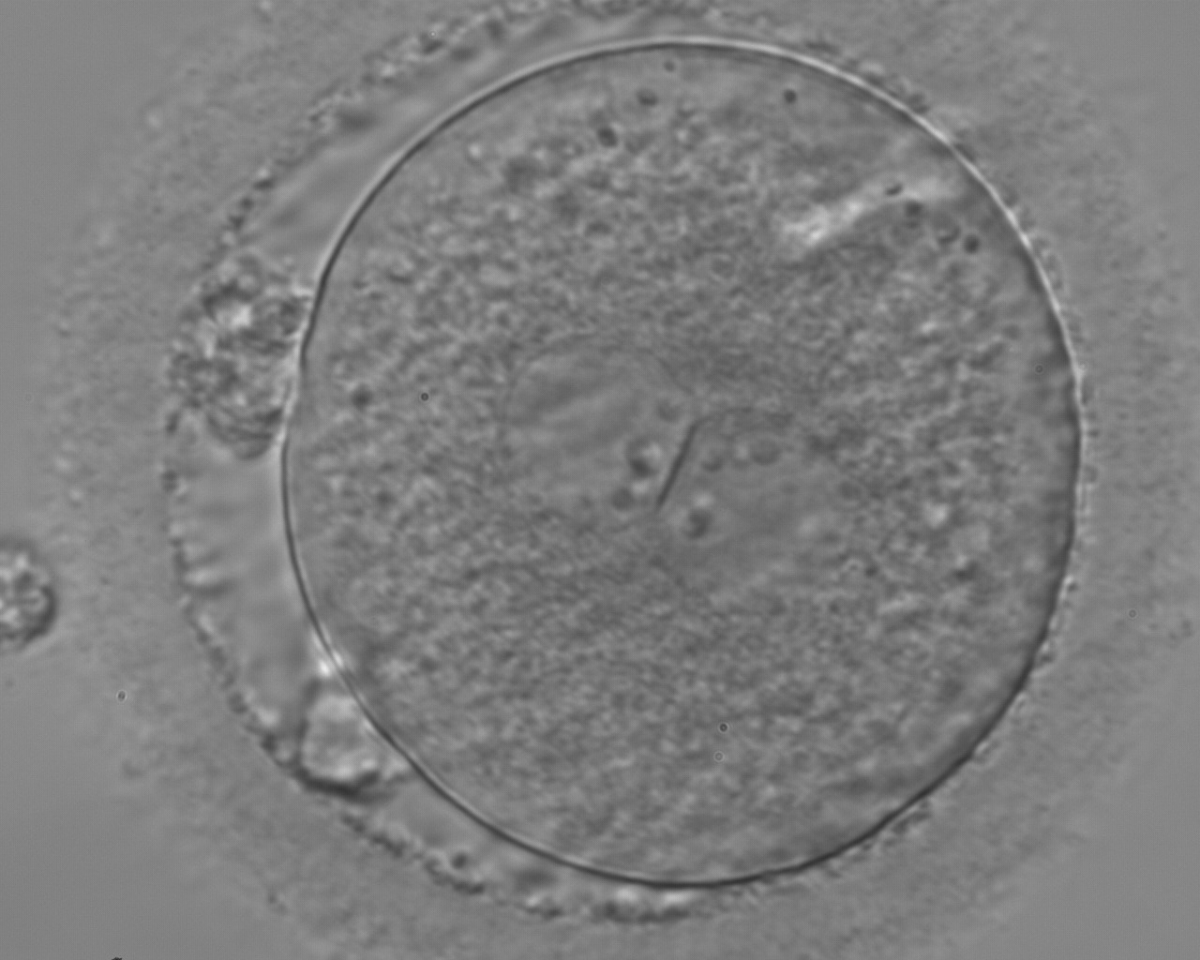
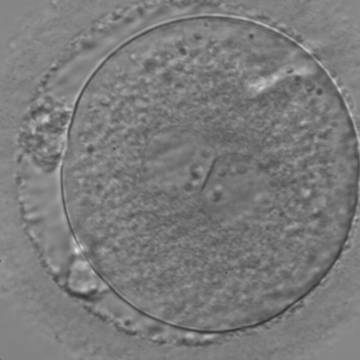
Figure 130
A zygote with two centrally located and perfectly juxtaposed PNs in a slightly granular cytoplasm with a clear cortical zone (400× magnification). There is debris in the PVS as well as fragmentation of one polar body (presumably the first polar body), which is significantly separated from the other polar body (presumably the second polar body). It was transferred but the outcome is unknown.
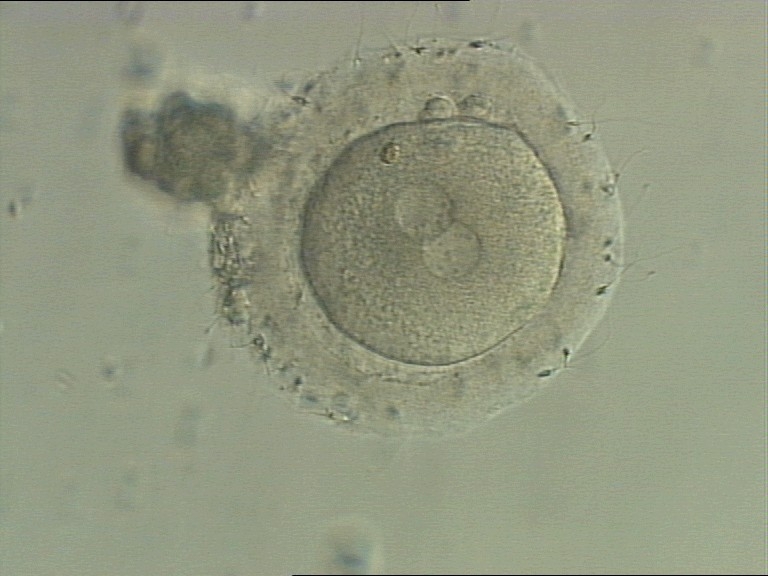
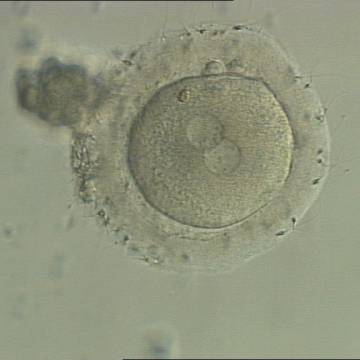
Figure 131
A zygote generated by IVF with a thick ZP (400× magnification). PNs are juxtaposed in the cytoplasm, which has a clear cortical zone. NPBs are small and aligned in one PN and scattered in the other. A refractile body is visible at the 11 o'clock position in this view.
Failed progression to apposition and syngamy (Figs 132–134) mostly depends on sperm centrosome activity. Observations of zygotes with separated PNs at fertilization check during subsequent development (Figs 135–136) demonstrate severe delay or arrest in development (Fig. 136) in almost 80% of cases (Gianaroli et al., 2003). This condition is most frequently associated with pathological spermatozoa, particularly from epididymal or testicular samples (Fig. 133).
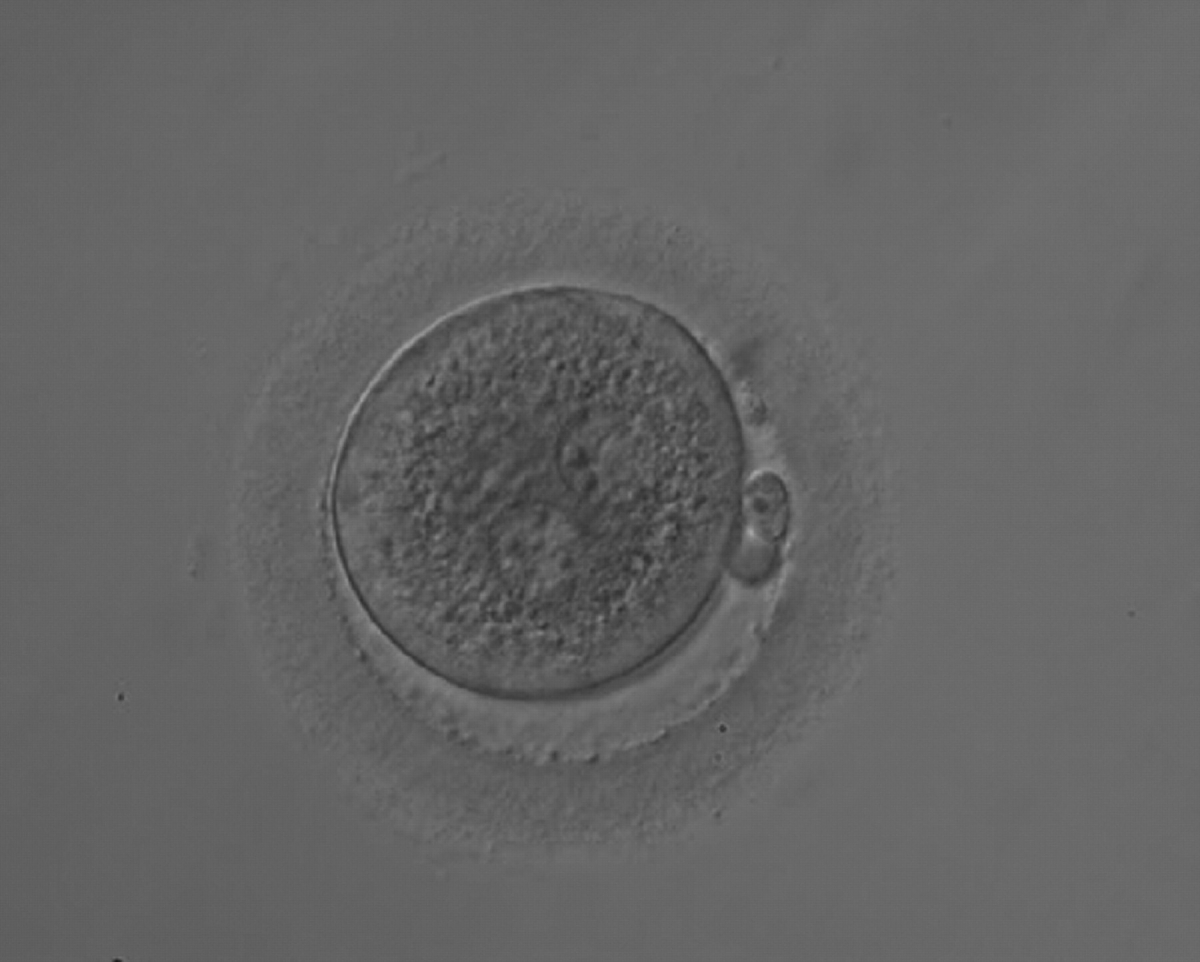
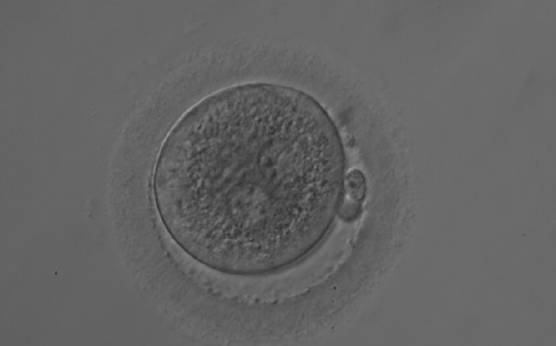
Figure 132
A zygote 18 h post-ICSI displaying two PNs that are not juxtaposed (200× magnification). NPBs are of different size, aligned in one PN and scattered in the other. The cytoplasm is very granular.
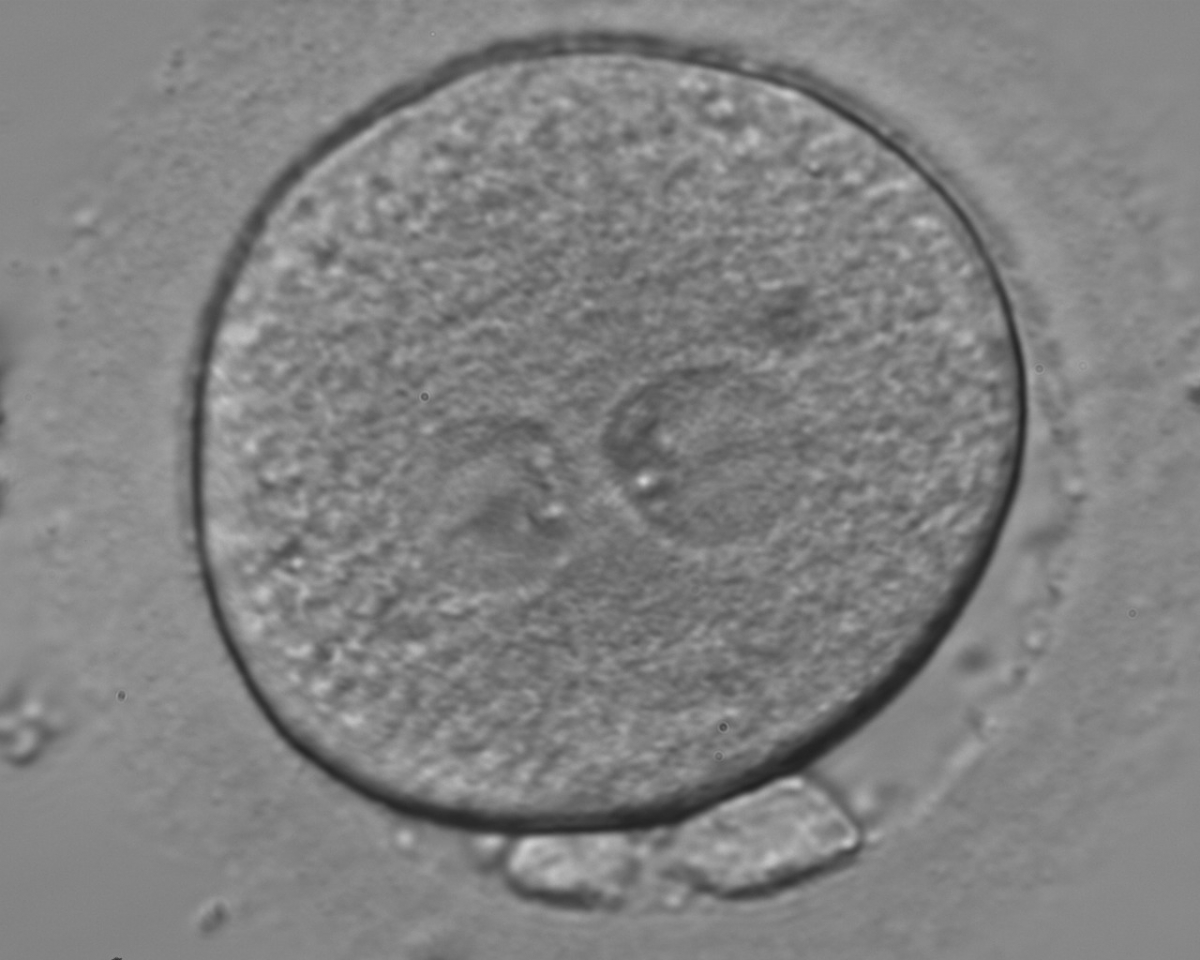
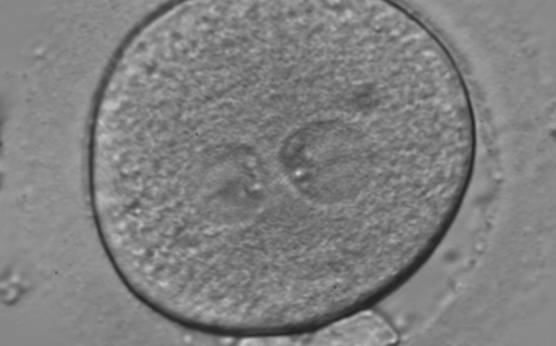
Figure 133
A zygote generated by ICSI using fresh epididymal sperm, observed 18 h post-insemination (400× magnification). PNs are not juxtaposed, different in size and NPBs are symmetrically distributed. It was transferred but failed to implant.
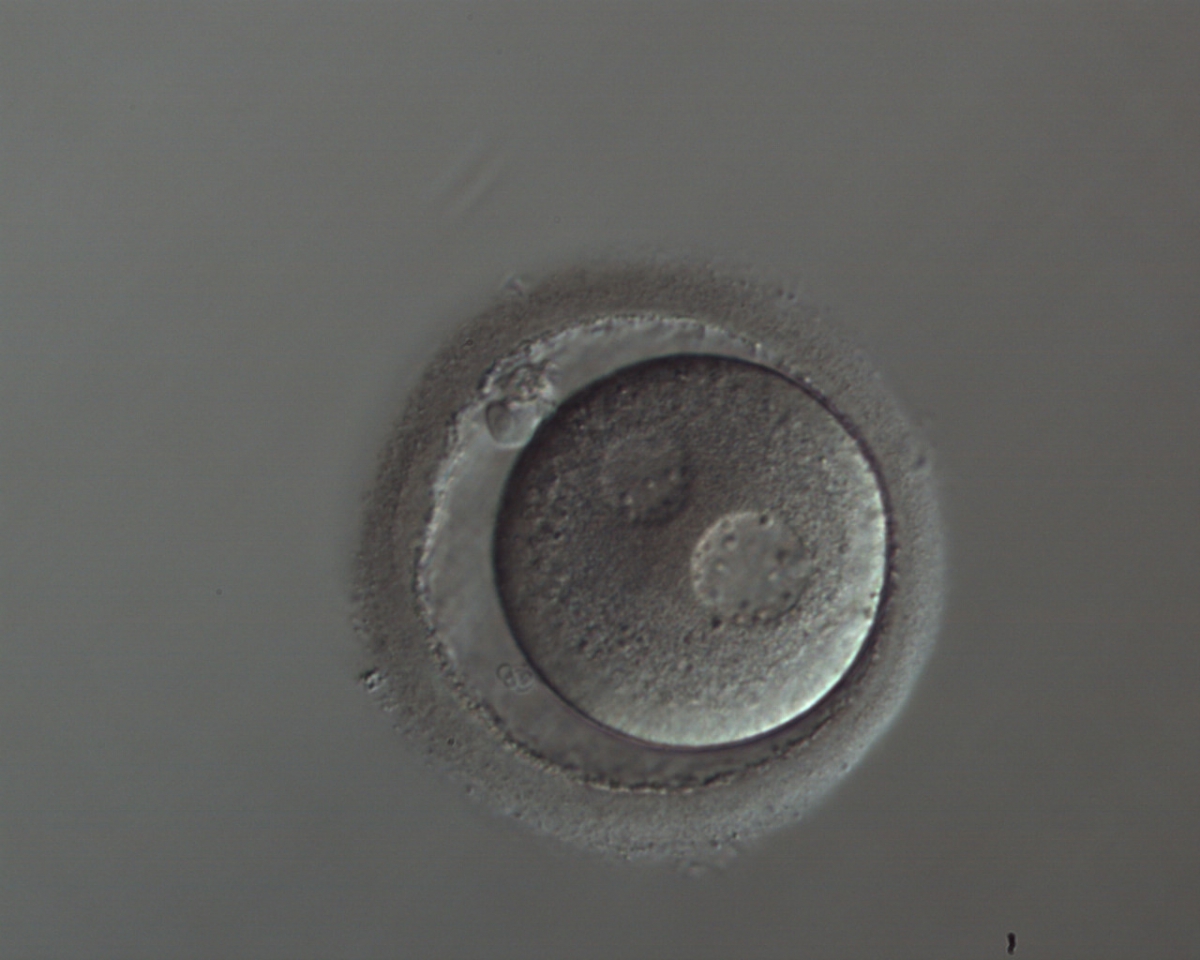
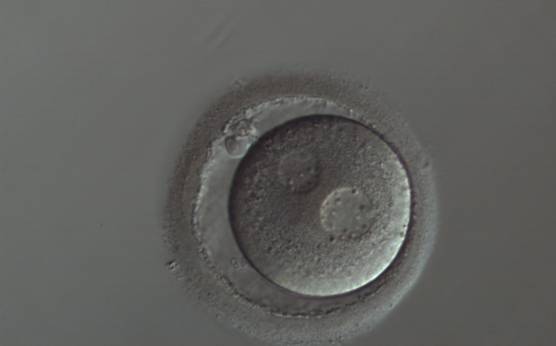
Figure 134
A zygote with widely separated and unequal-sized PNs, which show scattered small NPBs (150× magnification). The cytoplasm appears slightly granular and the PVS is enlarged. It was transferred but implantation outcome is unknown.
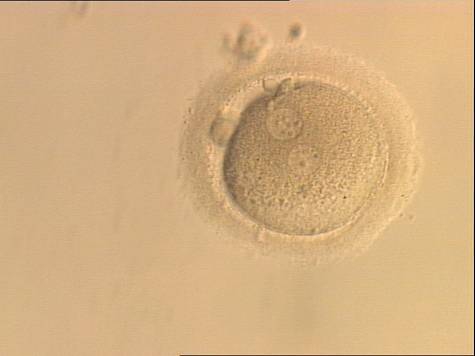
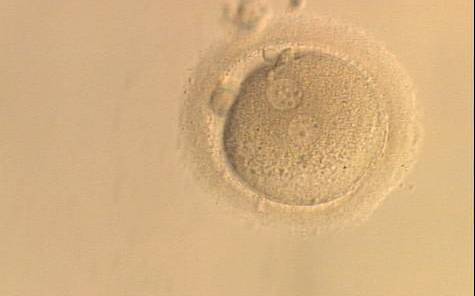
Figure 135
A zygote generated by ICSI and observed at 16, 17.5, 19 and 22 h after insemination (200× magnification). The two PNs are peripherally located, widely separated and have an unequal number of scattered NPBs. It was discarded.
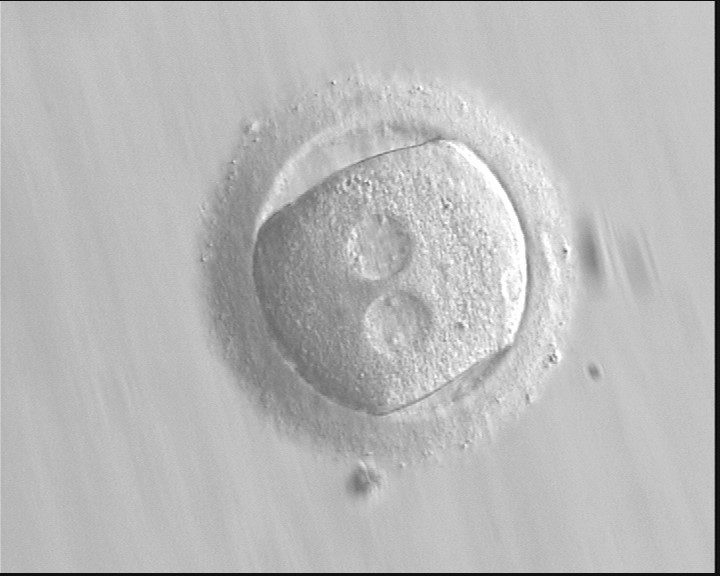
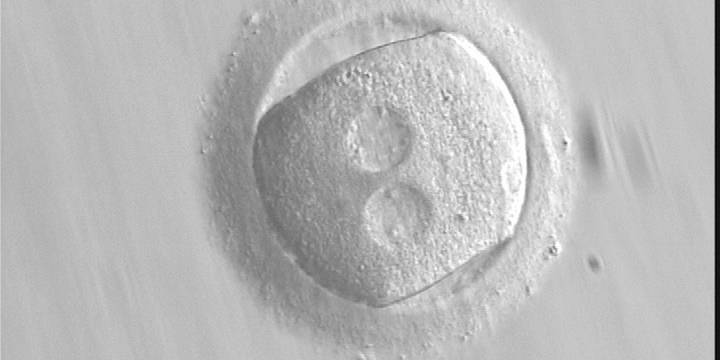
Figure 136 (a)
A zygote with an irregular shape, generated by ICSI and observed 18 h post-ICSI (400× magnification). The polar body had been biopsied. PNs are widely separated and contain scattered small NPBs.
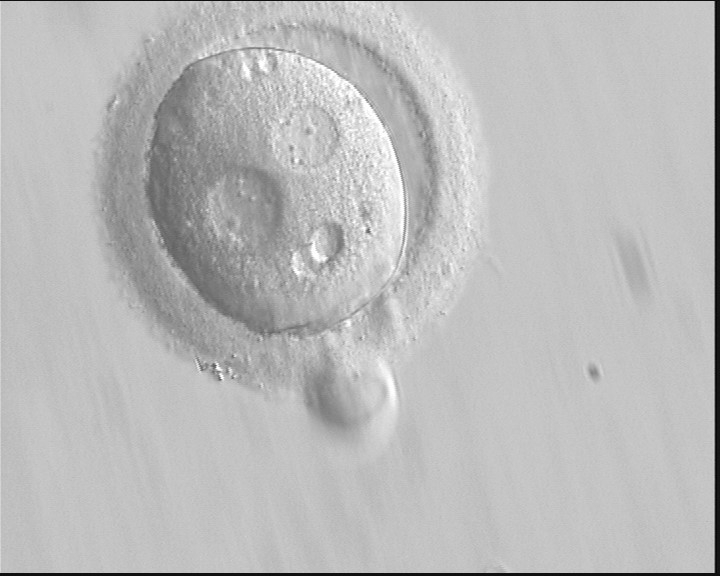
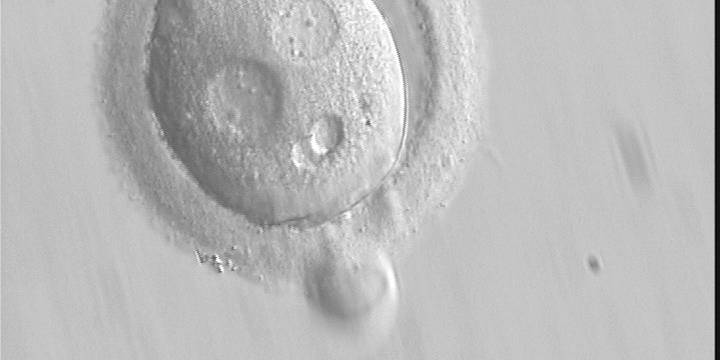
Figure 136 (b)
Observed 20 h post-ICSI (400× magnification). The cytoplasm is normal in colour but displays two vacuole-like structures. It failed to cleave during further development.
C.4 Centrally/peripherally positioned PNs
The position of PNs has a relevant effect on the first cleavage plane that normally goes through the pronuclear axis (Scott, 2003). In the majority of zygotes with centrally positioned PNs (Fig. 137), the first cleavage occurs regularly, giving rise to normally developing embryos. Due to the dynamics of PN movements within the cytoplasm, their orientation on the polar axis varies depending on the progression of rotation (Figs 138 and 139) towards the final state which determines the first cleavage plane (Fig. 140).

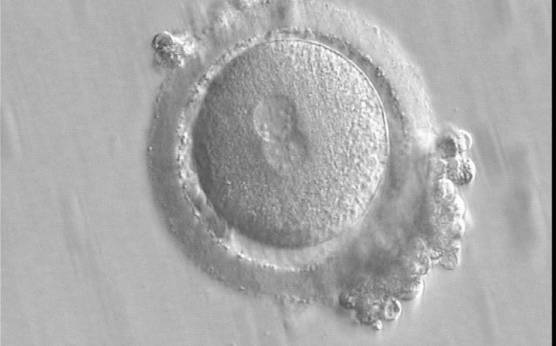
Figure 137
A zygote displaying centrally located, and in this view, partially overlapping PNs (400× magnification). The PN longitudinal axis is parallel to the polar bodies. NPBs are scattered in both PNs with respect to the PN junction but differ in number. It was transferred and implanted.
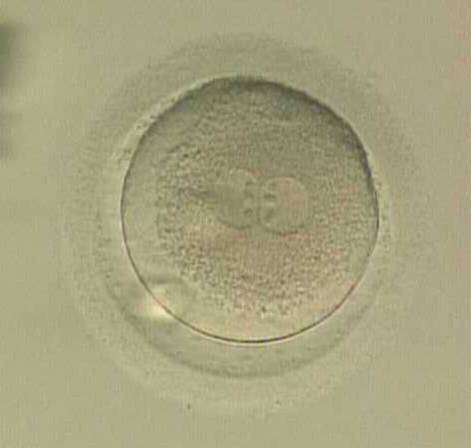
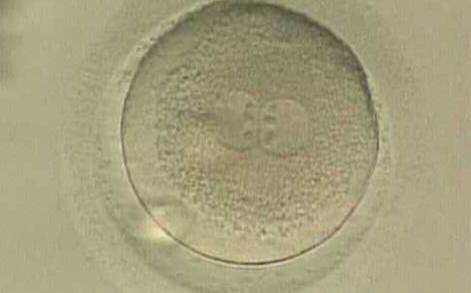
Figure 138
A zygote displaying centrally located PNs in which NPBs are symmetrically distributed and aligned with respect to the PN junction (400× magnification). A clear cortical zone, the halo, appears in the cytoplasm and there is a slightly enlarged PVS. It was transferred but implantation outcome is unknown.
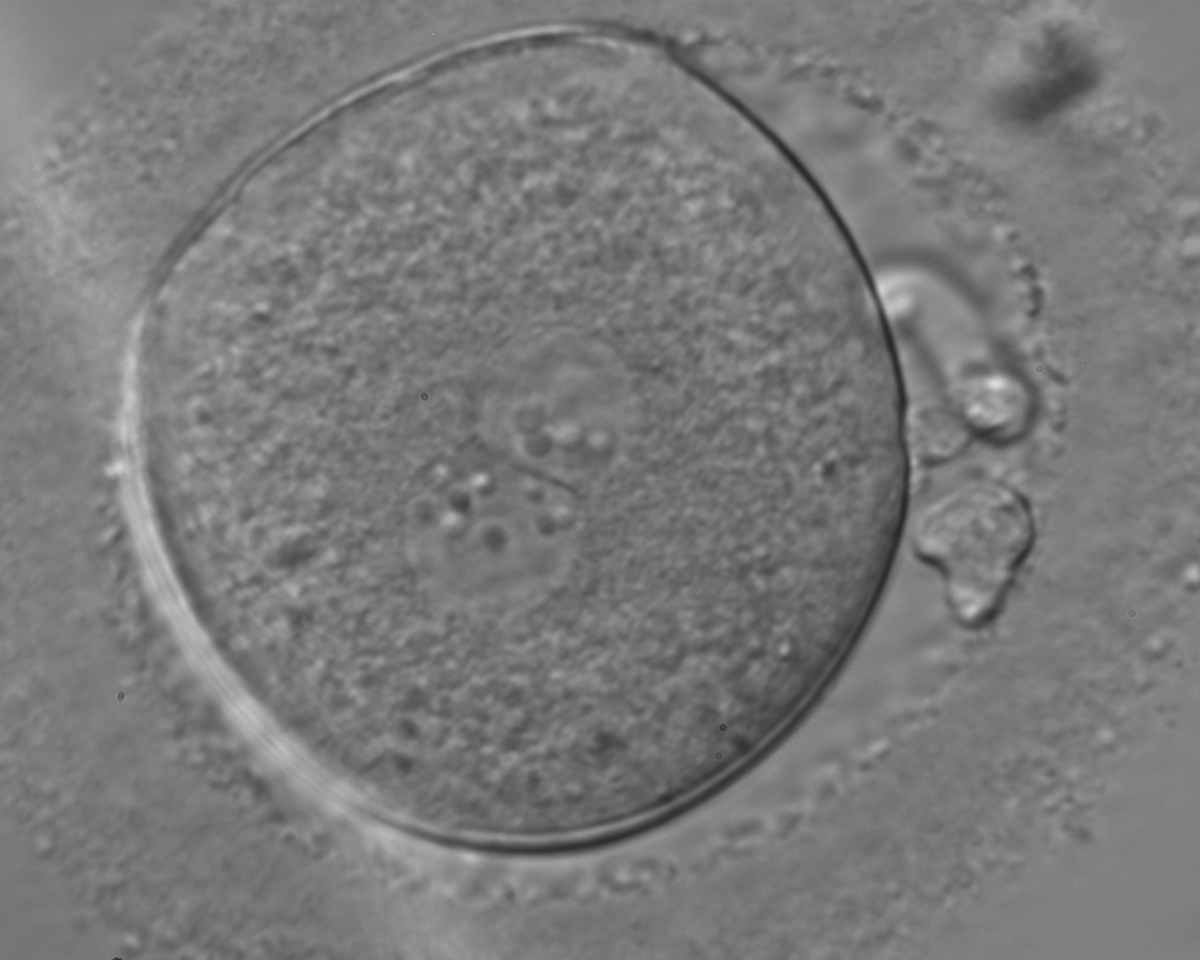
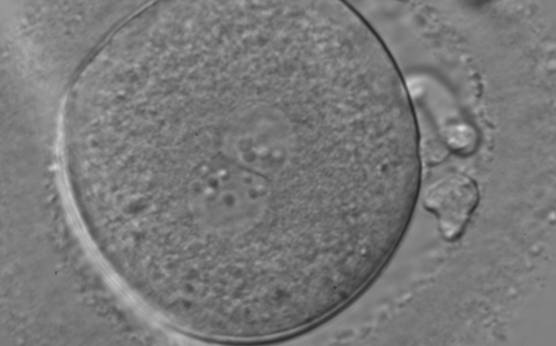
Figure 139
A zygote, generated by ICSI and observed 18 h post-insemination displaying centrally located PNs in which NPBs differ in number and size (400× magnification). The PN longitudinal axis is almost tangential to the plane through the polar bodies. NPBs are not symmetrically distributed, aligned in one PN and scattered in the other with respect to the PN junction. The ZP is thick and brush-like. It was transferred and failed to implant.
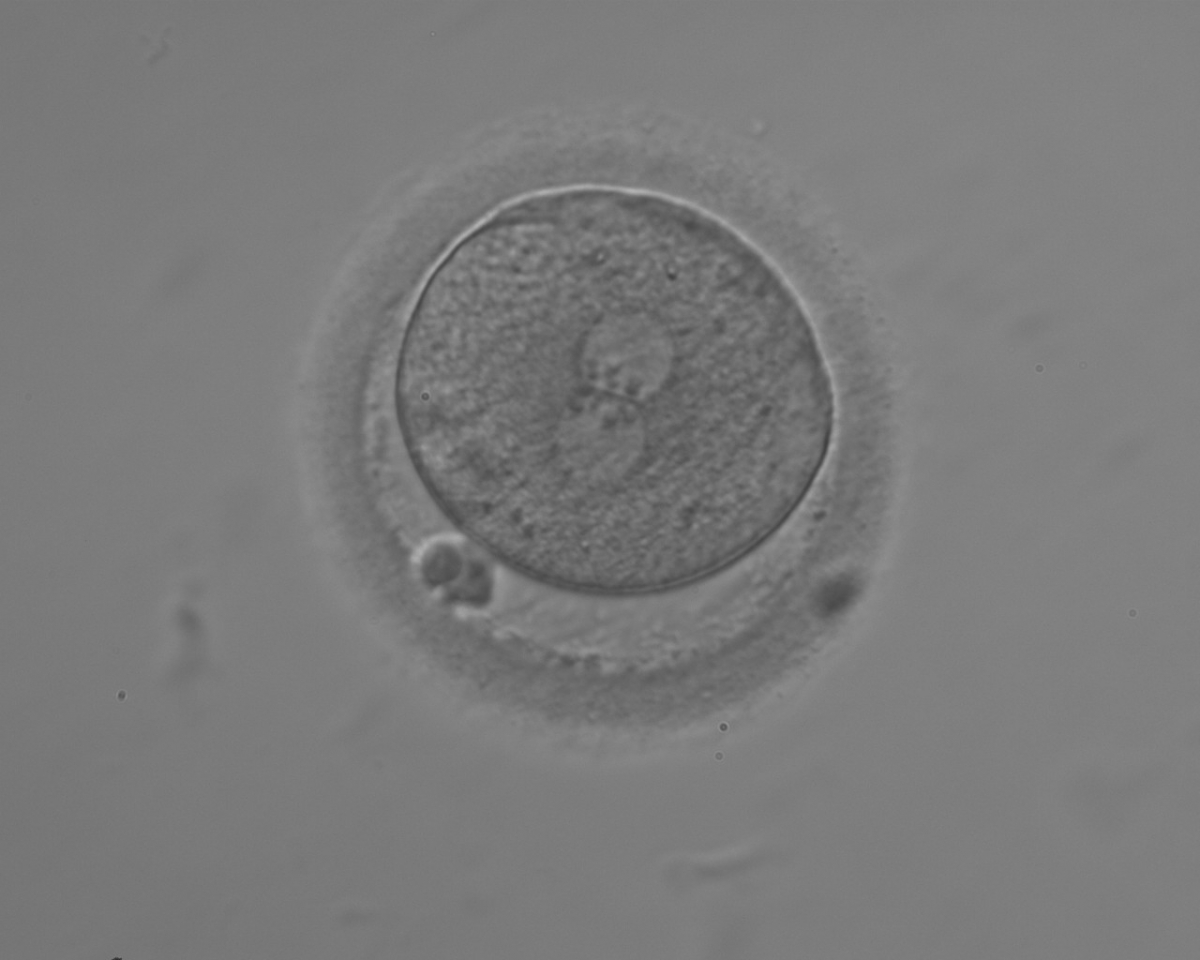
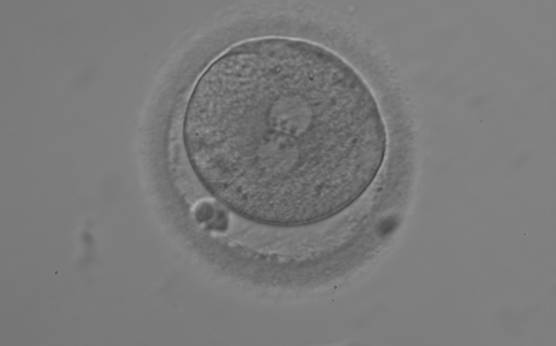
Figure 140
A zygote with equal numbers of large-sized NPBs aligned at the PN junction (200× magnification). PNs are juxtaposed and centralized; polar bodies are aligned tangential to the longitudinal axis through the PNs.
In cases of peripherally positioned PNs (Figs 141 and 142), cleavage occurs according to the pronuclear axis and results frequently in abnormal morphology (Fig. 143), cleavage (Fig. 144) and arrest of development (Fig. 145). Nevertheless implantation can occur (Fig. 146).
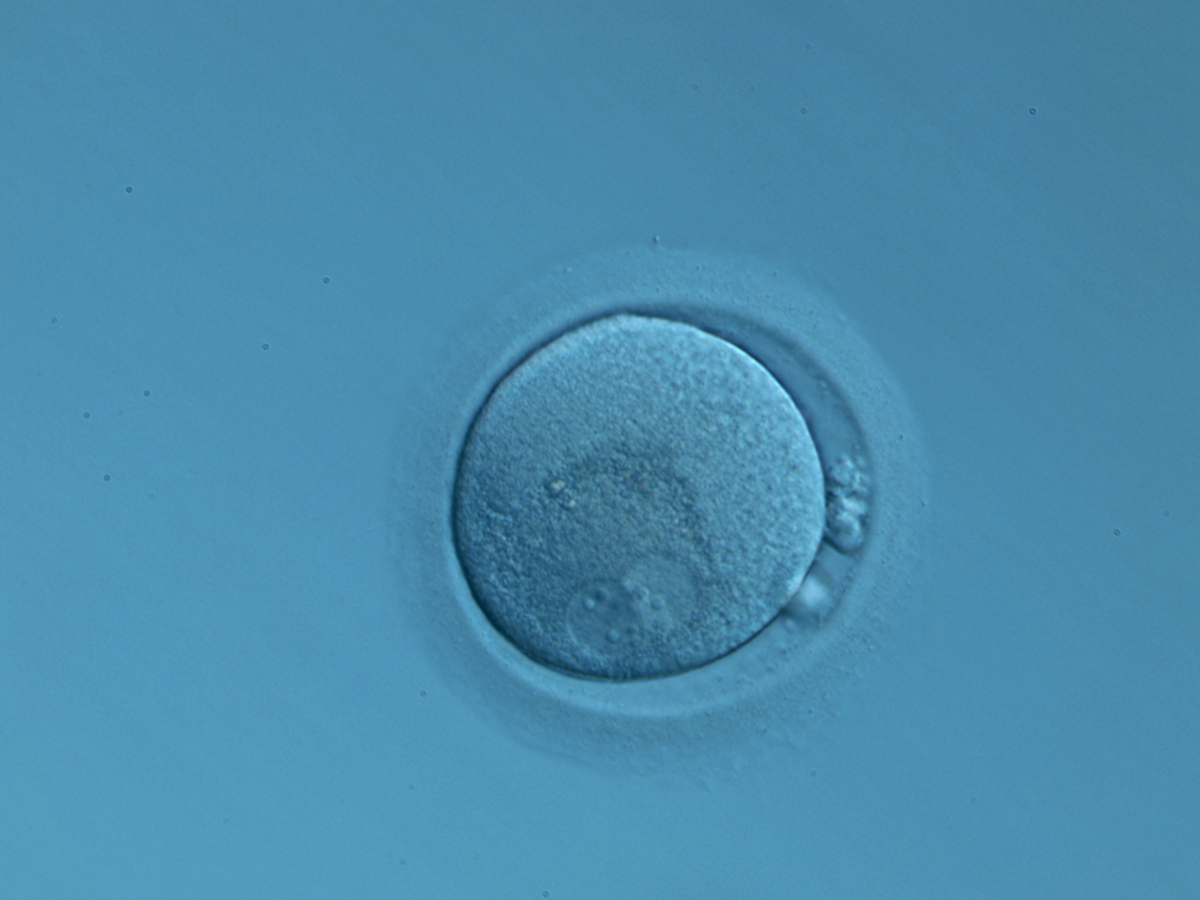
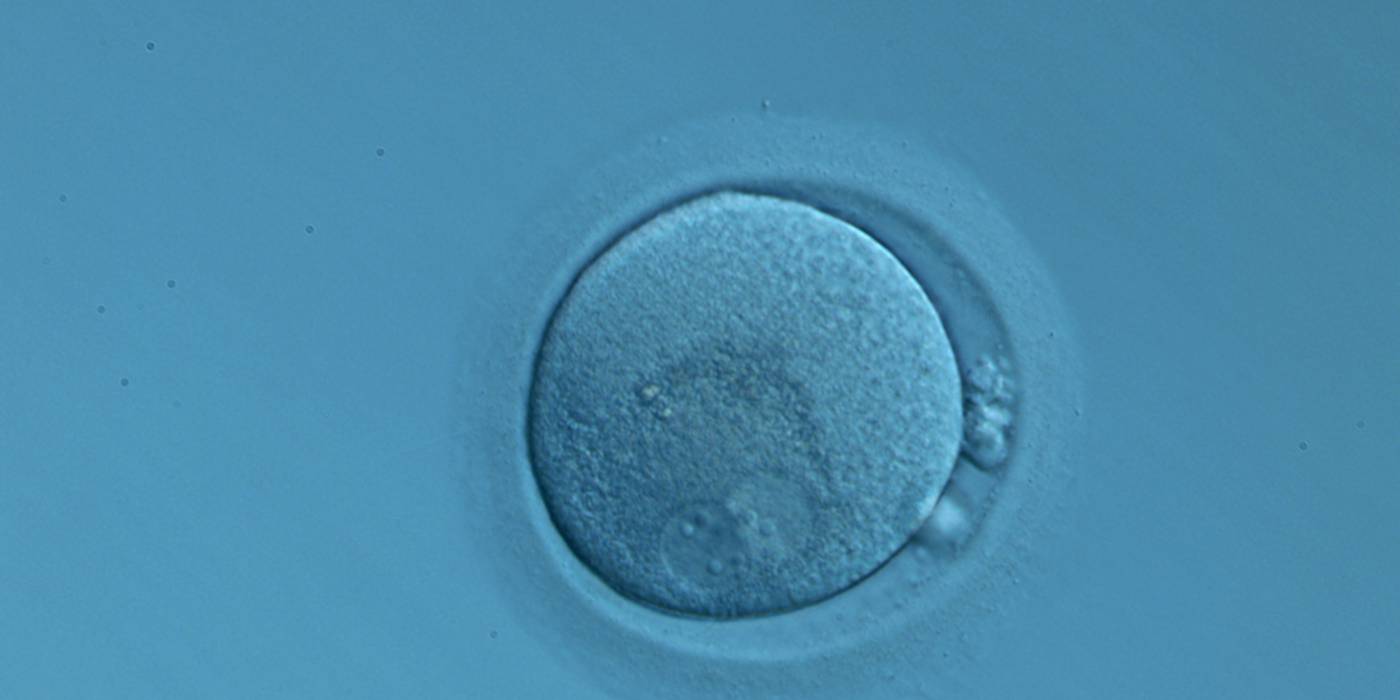
Figure 141
An ICSI zygote displaying two perfectly juxtaposed PNs in a peripheral position in the cytoplasm, which is showing central granularity due to clustering of the organelles (400× magnification). NPBs are different in number and distribution. One polar body (presumably the first polar body) is fragmented and the other is intact. It was transferred but the outcome is unknown.
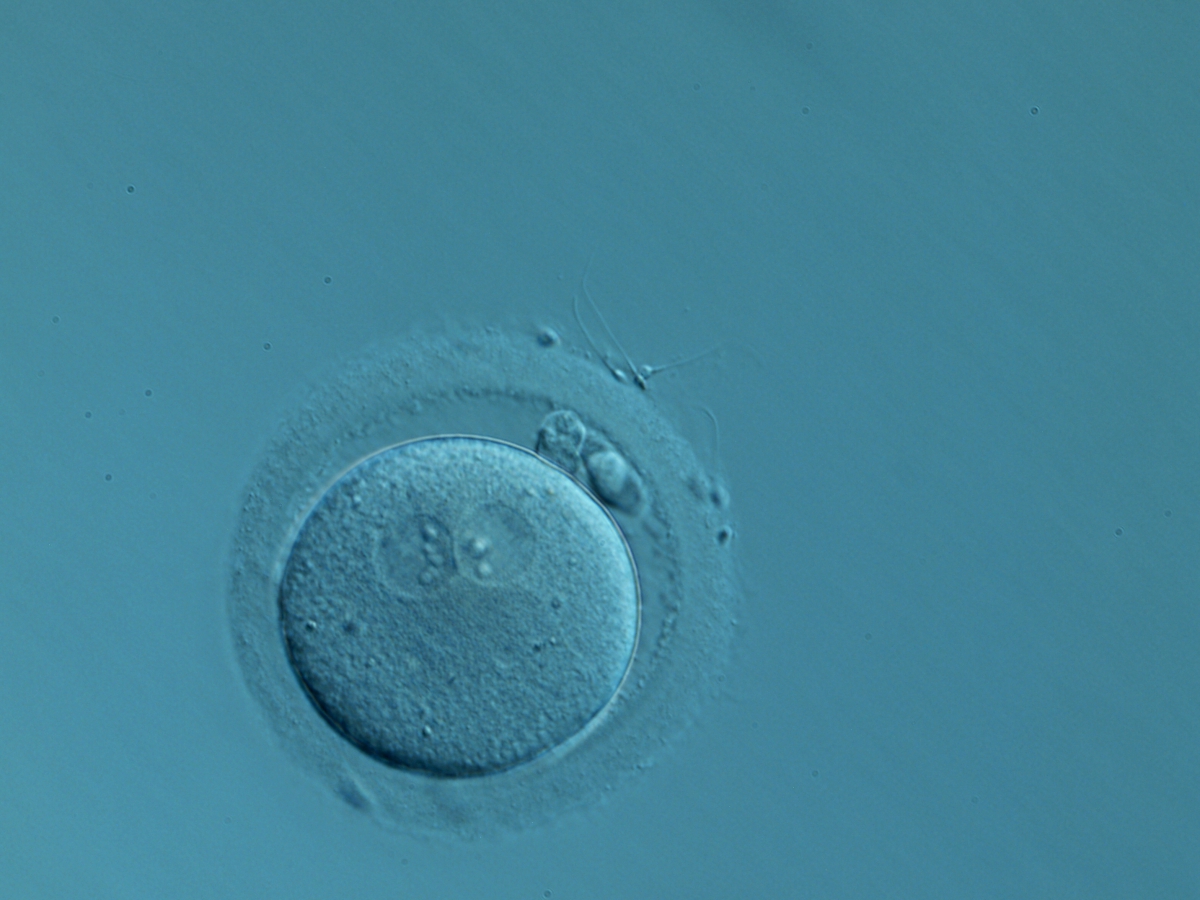
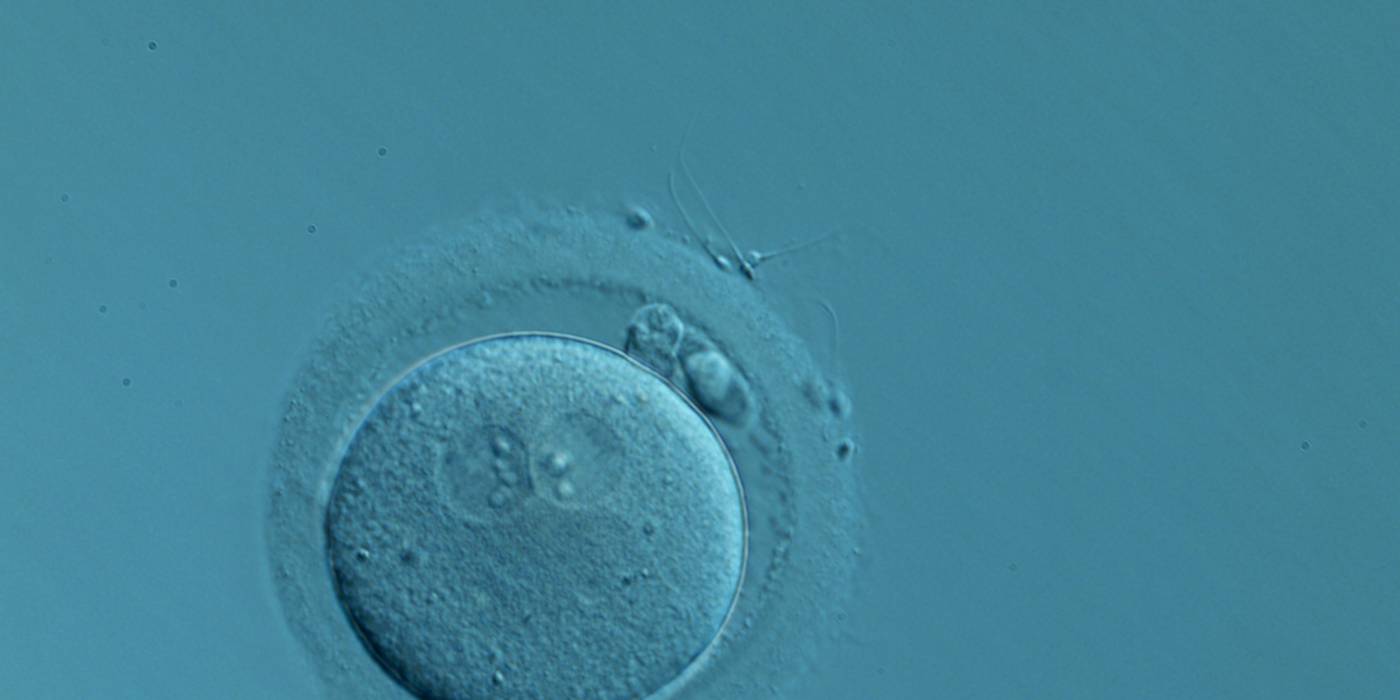
Figure 142
A zygote generated by IVF with normal-sized PNs slightly displaced to a peripheral position (400× magnification). NPBs are aligned and different in number. It was transferred but the outcome is unknown.
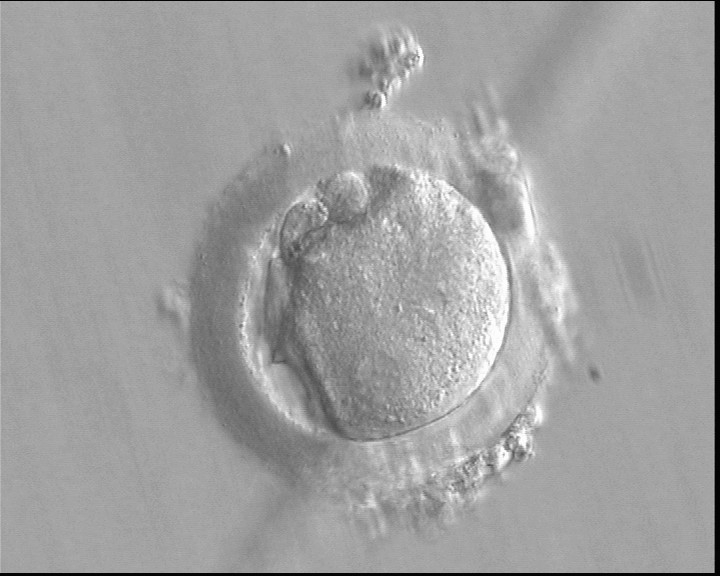
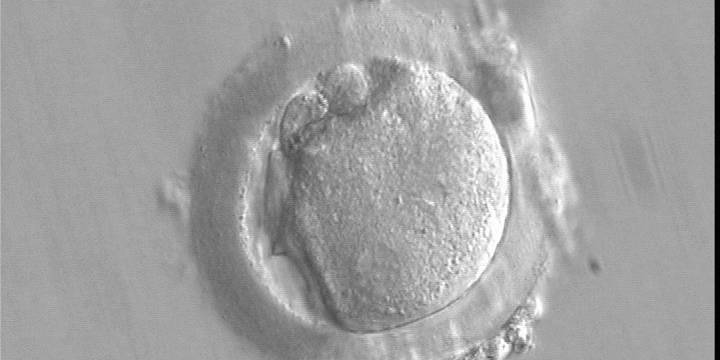
Figure 143
A deformed zygote at 16 h post-IVF displaying two PNs located peripherally in the cytoplasm (6–7 o'clock) that are parallel to the plane of the two large polar bodies (400× magnification).
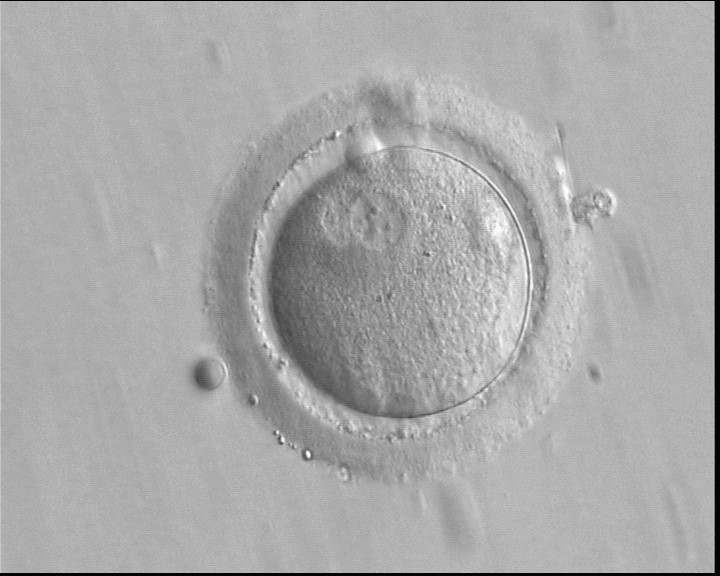
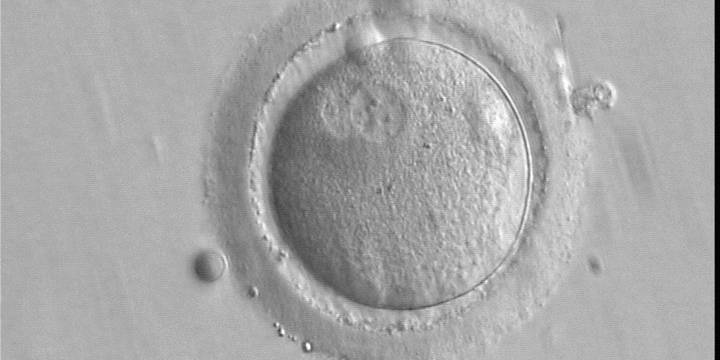
Figure 144
A zygote observed 16 h post-IVF showing peripheral PNs partly overlapping in this view (400× magnification). NPBs are small in size and scattered in both PNs. The derived embryo was highly fragmented with uneven blastomeres and was discarded.
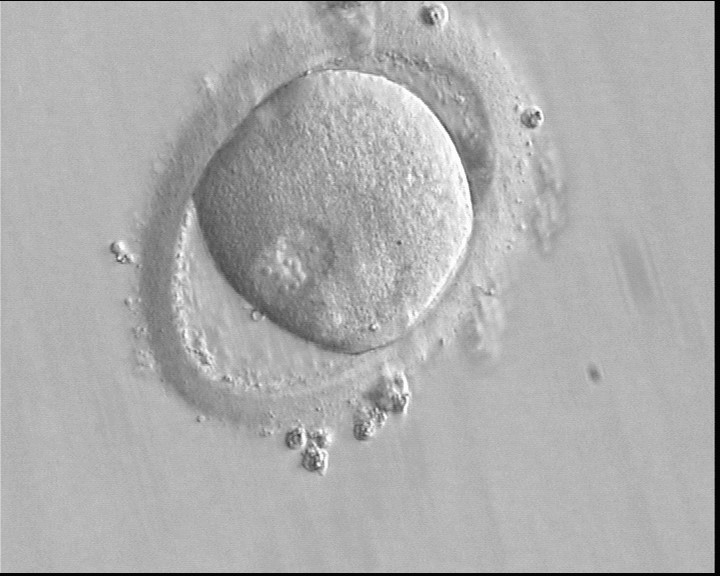
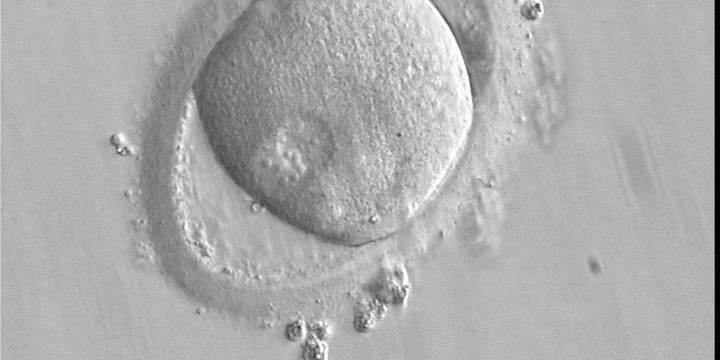
Figure 145
A zygote observed 16 h post-ICSI with peripherally positioned PNs (400× magnification). A large PVS is present inside a deformed, ovoid ZP. The oocyte has a clear cortical zone in the cytoplasm. It was not transferred because of arrested development.
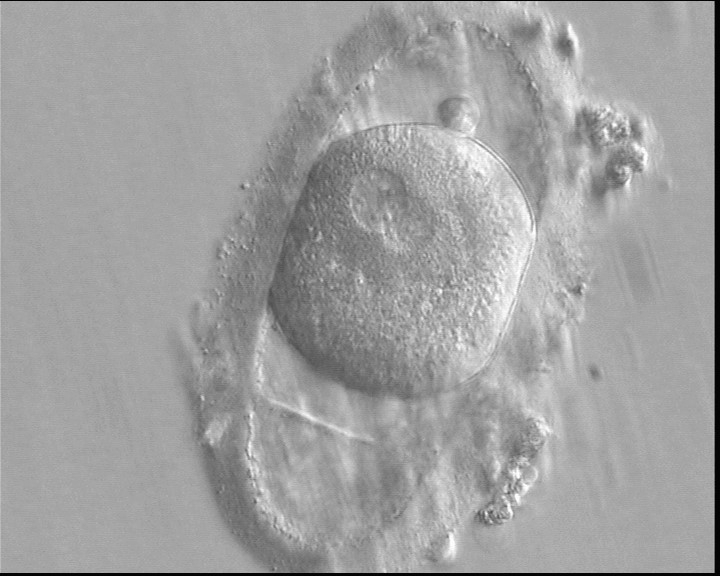
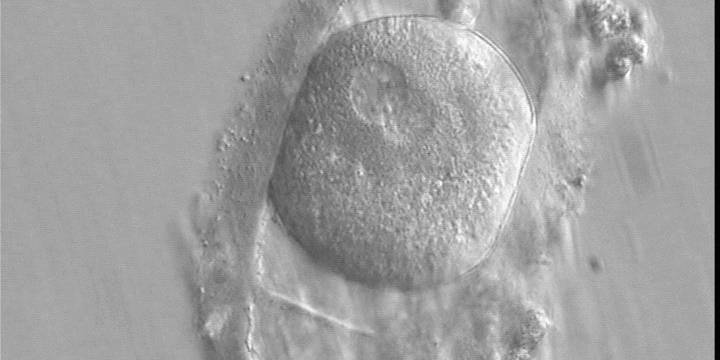
Figure 146
A zygote generated by ICSI displaying an ovoid ZP which demonstrates a duplication or tear in the layers (400× magnification). The fertilized oocyte is spherical and shows two PNs peripherally located and partly overlapping in this view. NPBs are small and scattered in both PNs. It was transferred and implanted.
C.5 Pronuclear membrane breakdown/syngamy
Following sperm entry, both PNs migrate toward each other while replicating their own DNA. At juxtaposition, nuclear membranes break down a few hours prior to initiation of the first cleavage division (Figs 147 and 148). Syngamy occurs (Fig. 149) and the two sets of haploid genomes merge.
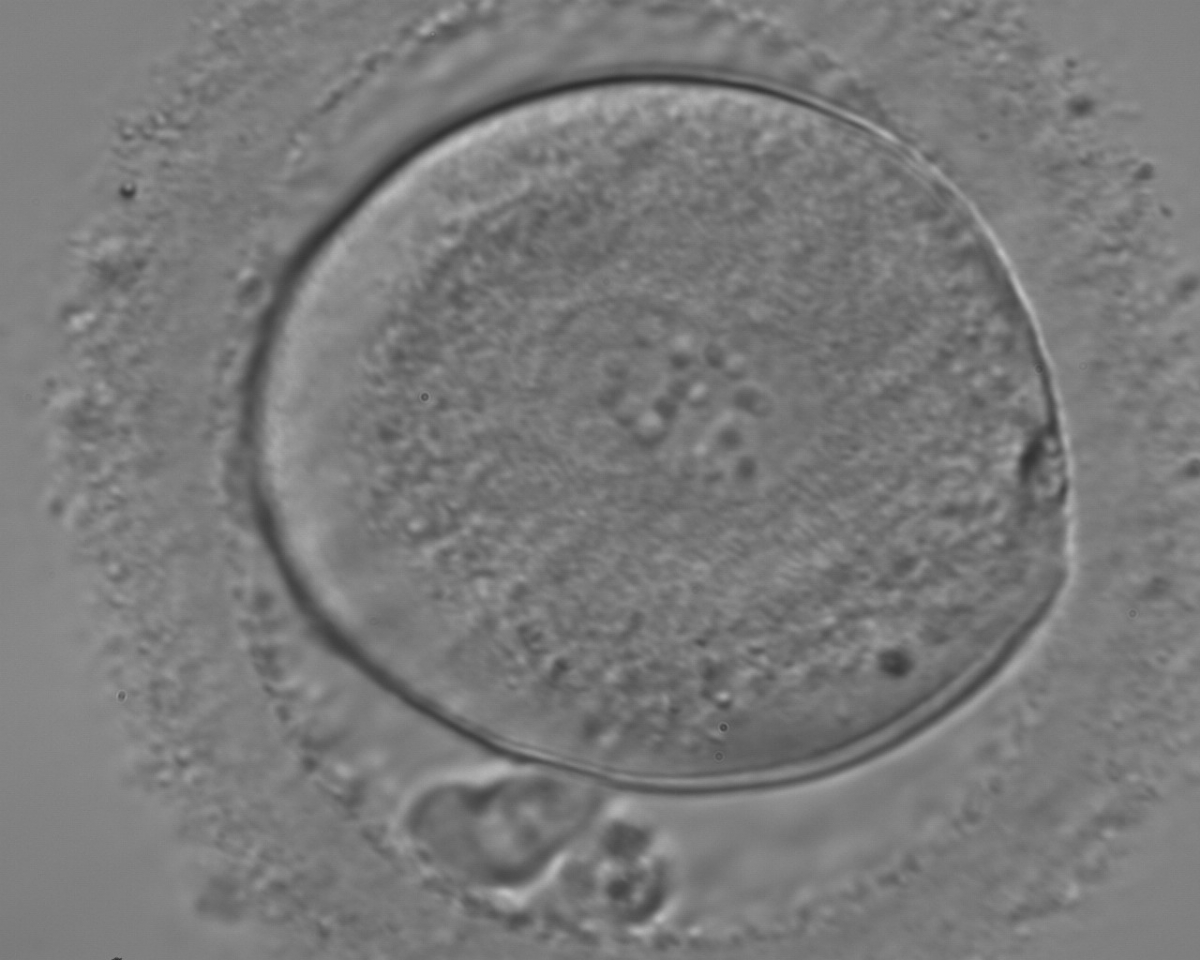
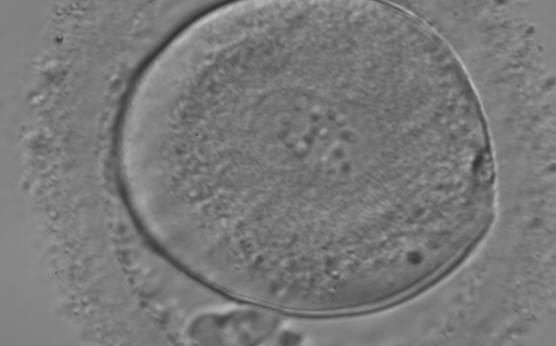
Figure 147
A zygote entering syngamy observed at 18 h after ICSI (400× magnification). The PN membranes are becoming indistinct and have large-sized NPBs. The ZP appears ‘brush-like’ and there is a clear cortical region evident in the peripheral cytoplasm. Further development resulted in cleavage to 4 cell and arrest on Day 3.
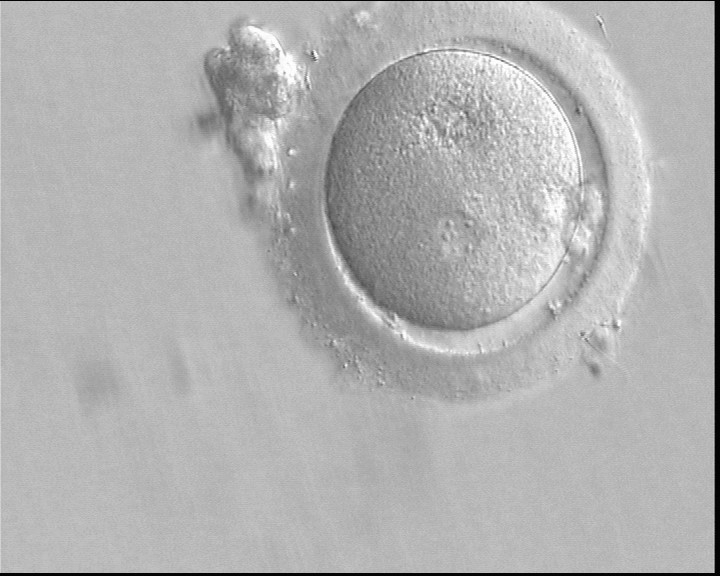
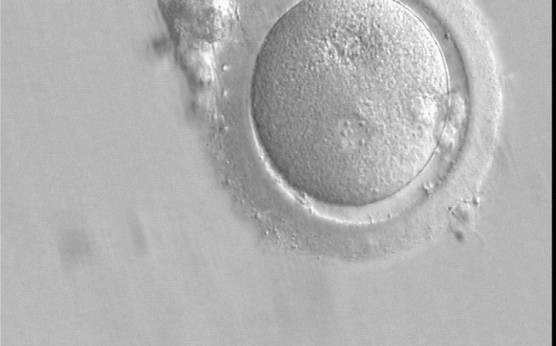
Figure 148
A zygote which is entering syngamy and is displaying membrane breakdown particularly evident in the upper PN (400× magnification). The observation was performed 15 h after ICSI. One of the two PNs and its associated NPBs are now very indistinct. It was transferred but failed to implant.
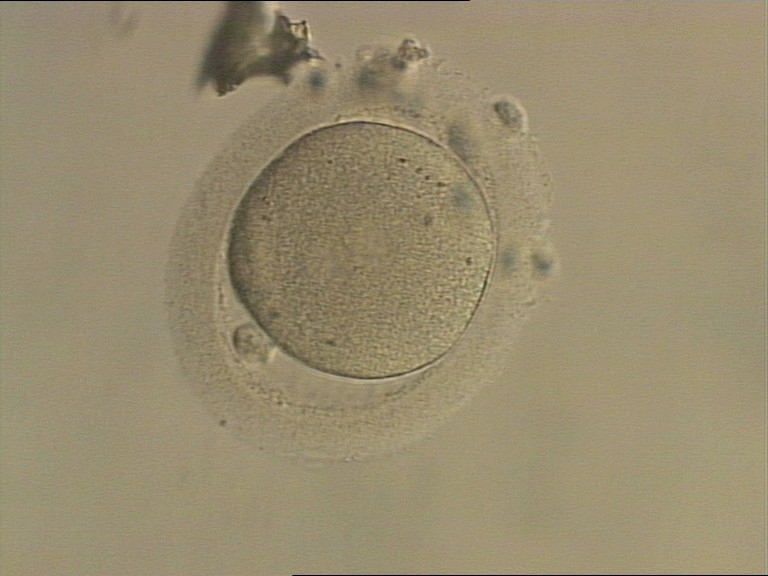
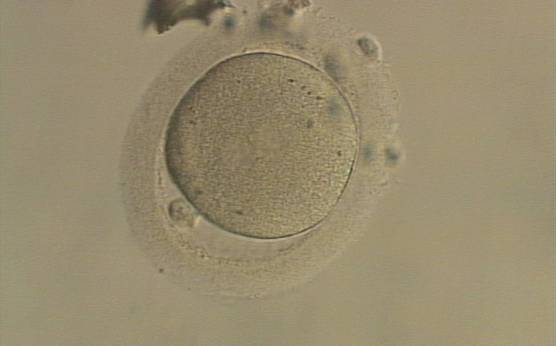
Figure 149
An ICSI zygote which has entered syngamy and is displaying pronuclear membrane breakdown (200× magnification). The NPBs are no longer distinct. It was transferred but the outcome is unknown.
The astral centrosome containing two centrioles splits and relocates to opposite poles of a bipolar spindle to establish bipolarization that controls cell division. The centrioles take a pivotal position on spindle poles, while chromosomes organize on the equator of the metaphase plate. Anaphase and telophase ensue completing the first mitotic division.
In contrast to some animal species, membrane fusion of PNs (Fig. 150) is not a common process in human zygotes.
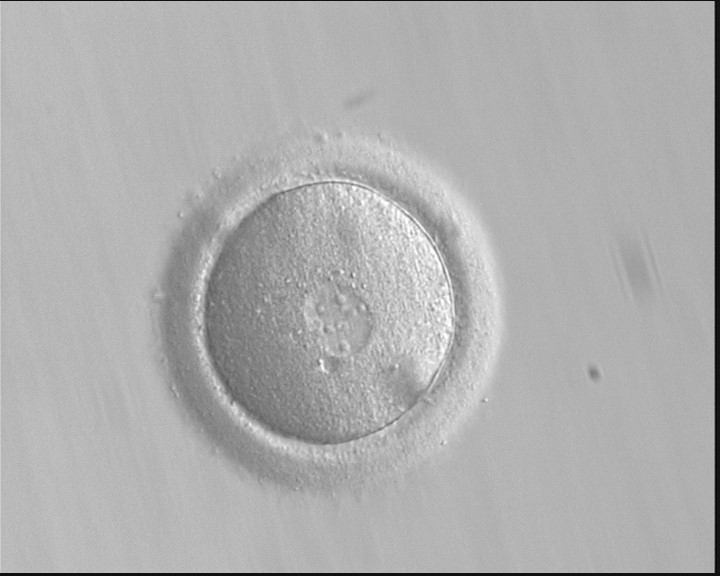
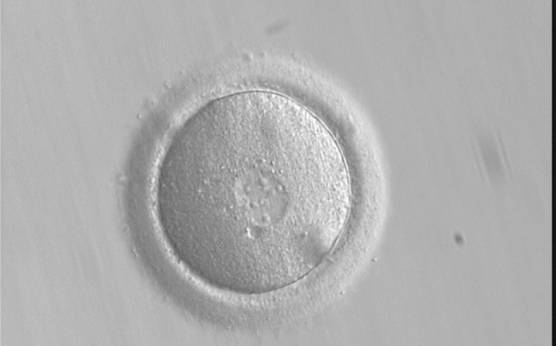
Figure 150
A zygote displaying PNs entering syngamy (400× magnification). The observation was performed 16 h after ICSI. The PN membranes are indistinct, particularly between the PNs where they have broken down, looking like PN membrane fusion. Large-sized NPBs are evident in the PNs. Polar bodies had been previously biopsied. It was transferred but failed to implant.
Article references:
Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum Reprod 2011;26:1270-1283.
Abstract/FREE Full Text
Edwards RG, Beard HK. Oocyte polarity and cell determination in early mammalian embryos. Mol Hum Reprod 1997;3:863-905.
Abstract/FREE Full Text
Gardner RL. Specification of embryonic axes begins before cleavage in normal mouse development. Development 2001;128:839-847.
Abstract
Garello C, Baker H, Rai J, Montgomery S, Wilson P, Kennedy CR, Hartshorne GM. Pronuclear orientation, polar body placement, and embryo quality after intracytoplasmic sperm injection and in vitro fertilization: further evidence for polarity in human oocytes? Hum Reprod 1999;14:2588-2595.
Abstract/FREE Full Text
Gianaroli L, Magli MC, Ferraretti AP, Fortini D, Grieco N. Pronuclear morphology and chromosomal abnormalities as scoring criteria for embryo selection. Fertil Steril 2003;80:341-349.
CrossRef | Medline | Web of Science | Google Scholar
Montag M, Liebenthron J, Köster M. Which morphological scoring system is relevant in human embryo development? Placenta 2011;32:S252-S256.
CrossRef | Medline | Web of Science | Google Scholar
Payne D, Flaherty SP, Barry MF, Matthews CD. Preliminary observations on polar body extrusion and pronuclear formation in human oocytes using time-lapse video cinematography. Hum Reprod 1997;12:532-541.
Abstract/FREE Full Text
Payne JF, Raburn DJ, Couchman GM, Price TM, Jamison MG, Walmer DK. Relationship between pre-embryo pronuclear morphology (zygote score) and standard day 2 or 3 embryo morphology with regard to assisted reproductive technique outcomes. Fertil Steril 2005;84:900-909.
CrossRef | Medline | Web of Science | Google Scholar
Scott L. Oocyte and embryo polarity. Semin Reprod Med 2001;18:171-183.
Google Scholar
Scott L. Pronuclear scoring as a predictor of embryo development. Reprod Biomed Online 2003;6:201-214.
CrossRef | Medline | Google Scholar
Senn A, Urner F, Chanson A, Primi MP, Wirthner DW, Germond M. Morphological scoring of human pronuclear zygotes for prediction of pregnancy outcome. Hum Reprod 2006;21:234-239.
Abstract/FREE Full Text
