C. Trophectoderm morphology
The TE cells can be clearly distinguished from the ICM cells as the blastocyst begins to expand (i.e. expansion Grade 3 or higher). The role of the TE cells in the early stages of blastocyst development is not entirely clear but their role in creating the fluid filled blastocoel may be a key parameter in ICM determination. The role of the TE cells, however, is better understood during and after implantation as they play a key role in apposition, adhesion and invasion of the endometrium, thus allowing the blastocyst to embed in the uterus. The TE cells also produce several molecular factors that aid in the implantation process (Aplin, 2000). Without properly functional TE cells, the embryo would remain within the ZP as these cells are actively involved in breaking free of the ZP (Sathananthan et al., 2003). The ultimate fate of the TE cells is to become the fetal extra-embryonic membranes as well as the placenta.
The TE cells have traditionally been scored in a similar manner to the ICM, i.e. by their number and cohesiveness according to three different grades (1–3). The best TE category (1) contains many cells that form a cohesive epithelium (Figs 365–368), the middle TE category (2) is composed of few cells forming a loose epithelium (Figs 369–372) and the worst category (3) describes a TE that contains very few, large cells that struggle to form a cohesive epithelium (Figs 373–376). The grading of the TE cells has been demonstrated in some reports to have an association with implantation (Zaninovic et al., 2001; Ahlström et al., 2011), whereas other publications have found no relationship between TE grade and viability (Richter et al., 2001).
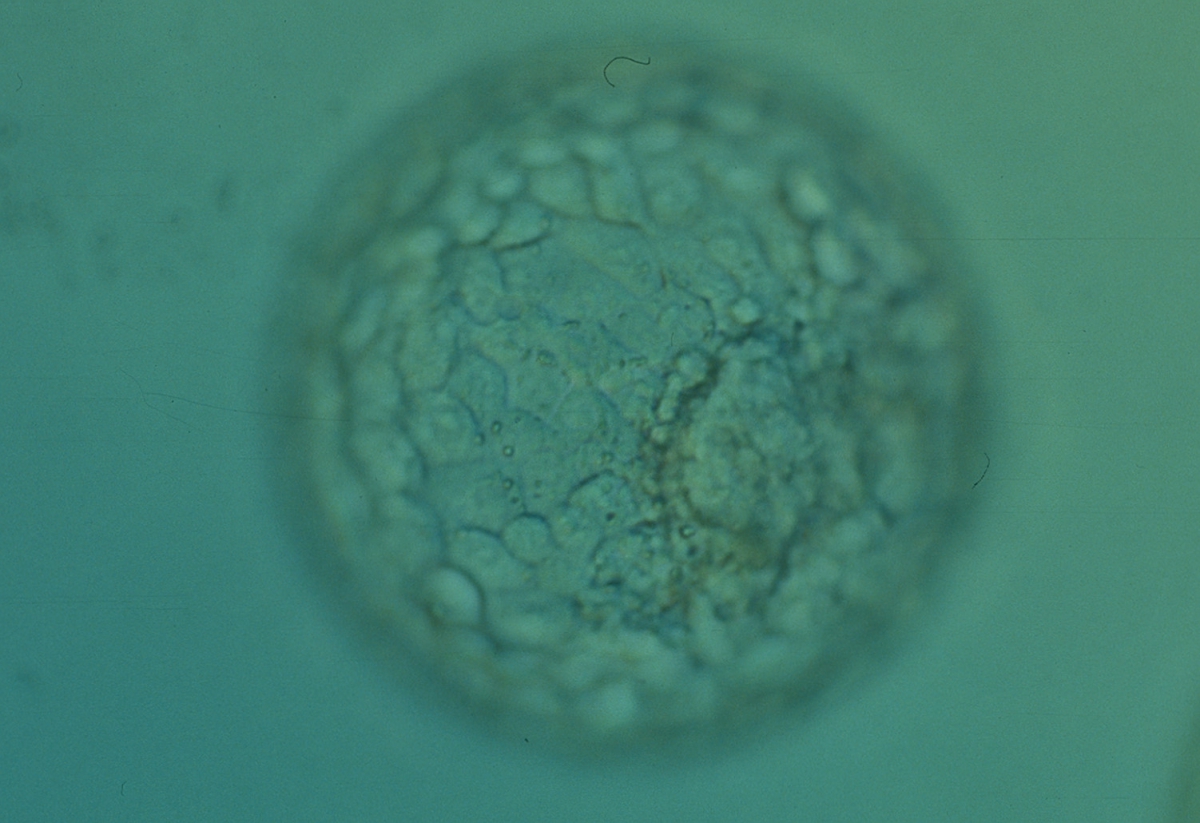
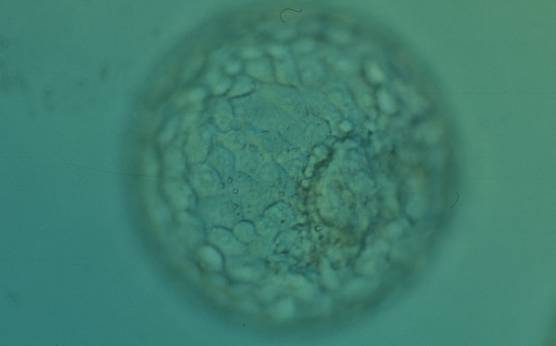
Figure 365
Expanded blastocyst (Grade 4:1:1) with many evenly sized cells making up a cohesive TE surrounding the enlarged blastocoel cavity. The ICM is clearly visible at the 4 o′clock position in this view. The blastocyst was transferred but the outcome is unknown.
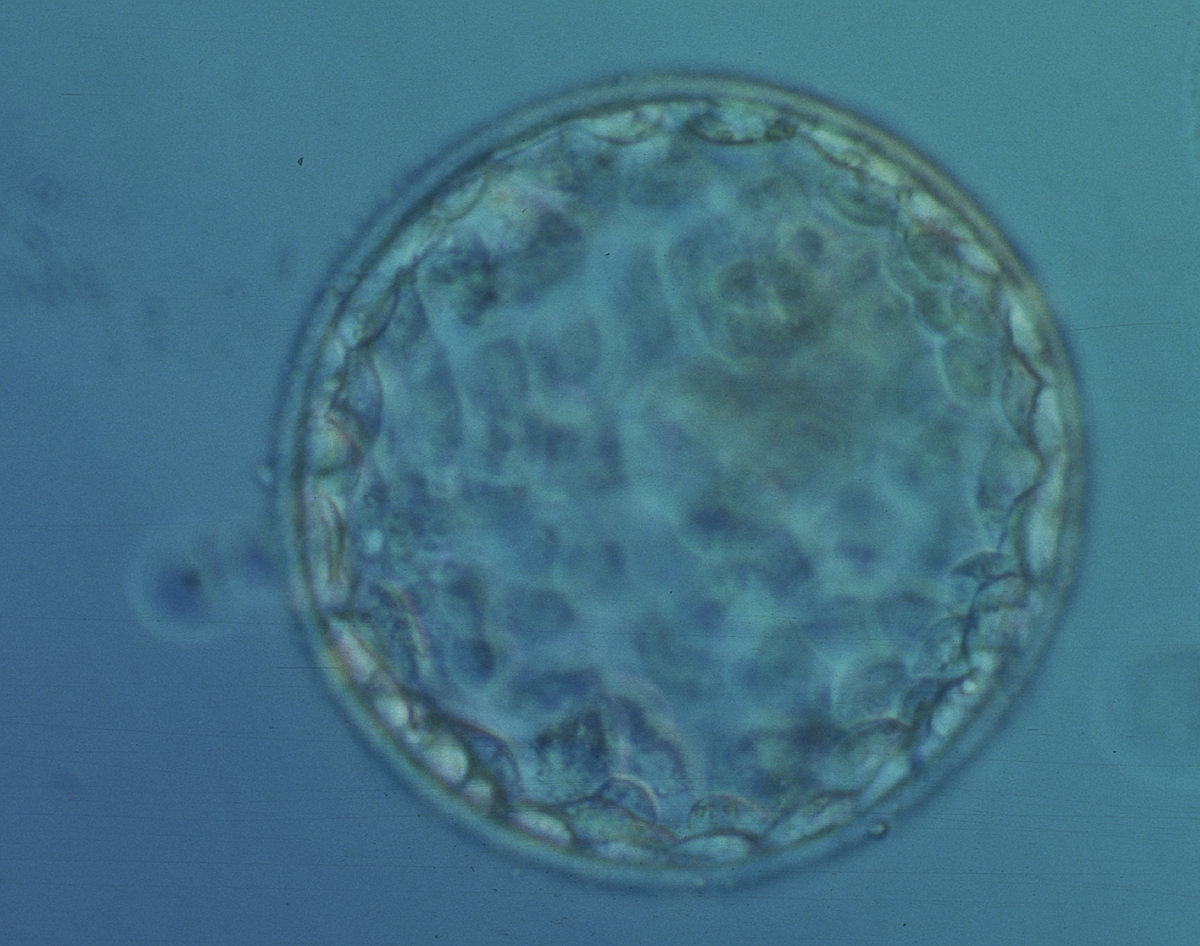
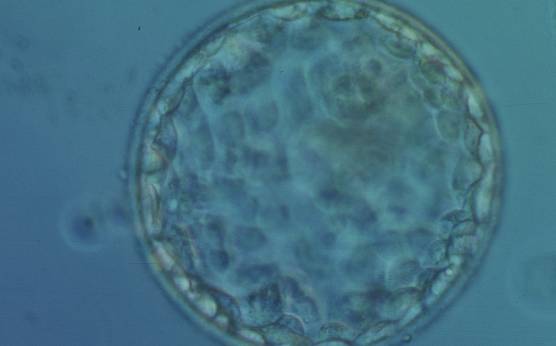
Figure 366
Early hatching blastocyst (Grade 5:1:1) with many cells making up a cohesive epithelium. The ICM is not clearly visible in this view. The blastocyst was transferred but failed to implant.
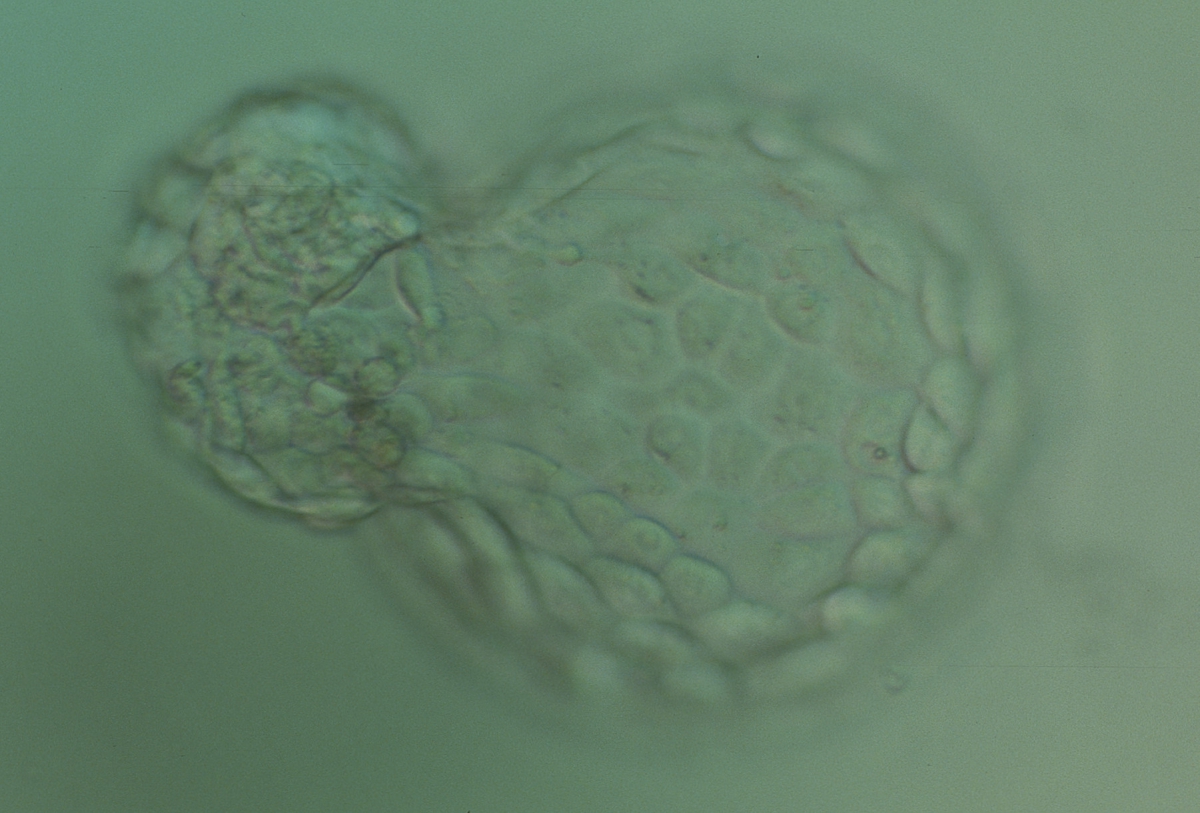
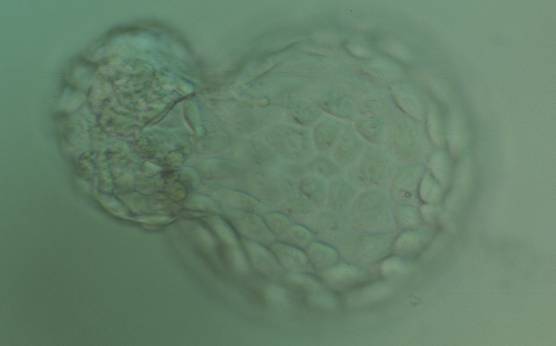
Figure 367
Hatching blastocyst (Grade 5:1:1) with many cells making up a cohesive TE. The ICM is herniating through a breach in the ZP at the 10 o′clock position in this view. The blastocyst was transferred but failed to implant.
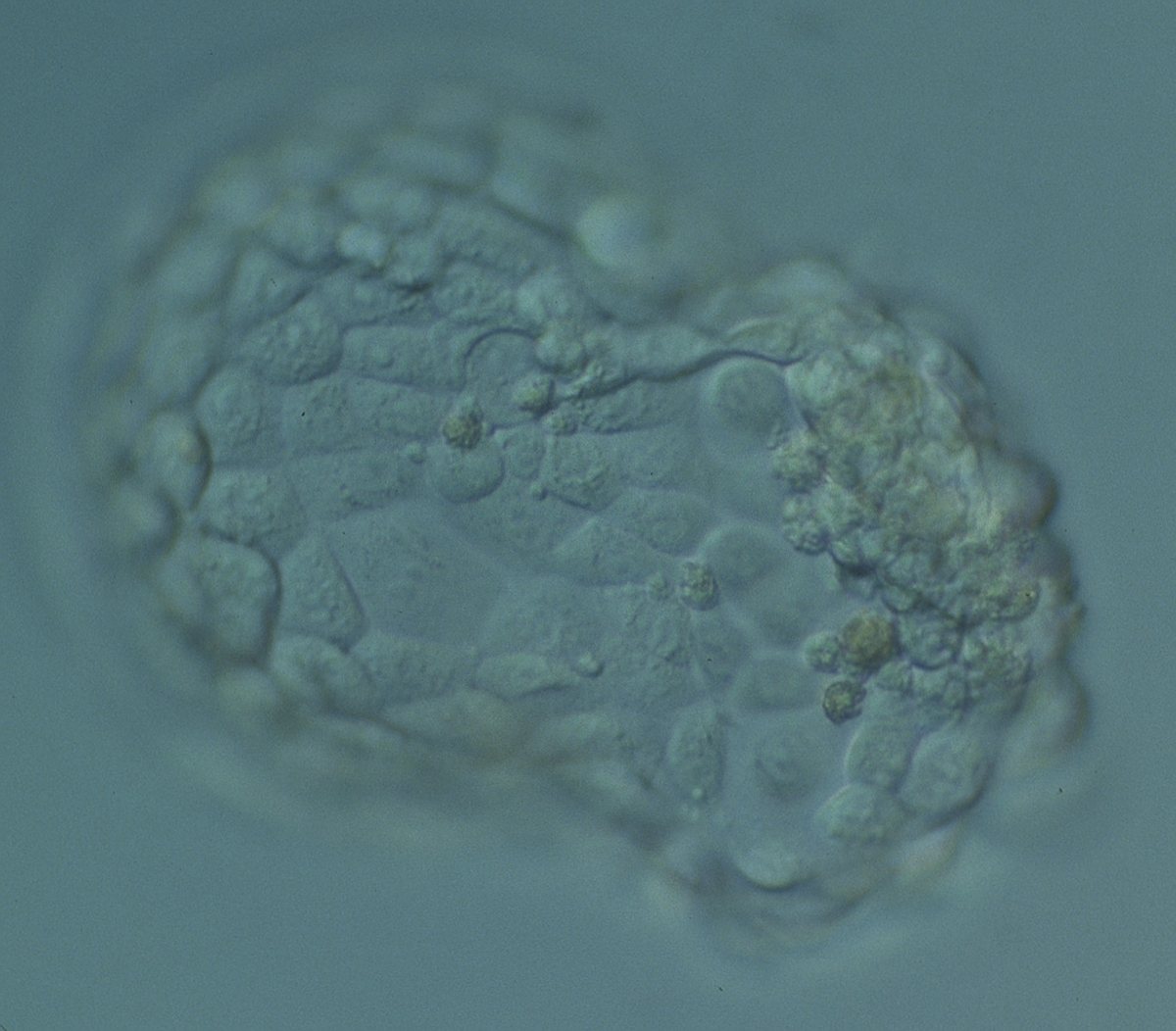
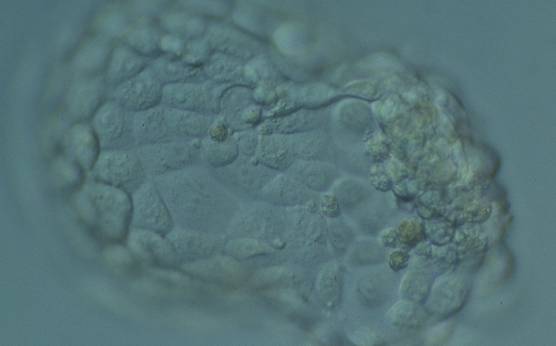
Figure 368
Hatching blastocyst (Grade 5:1:1) with many cells, some of variable size, making up a cohesive epithelium. The blastocyst was transferred but failed to implant.
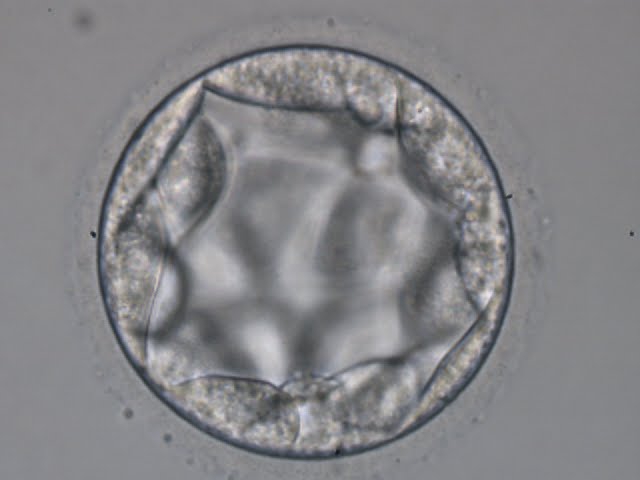
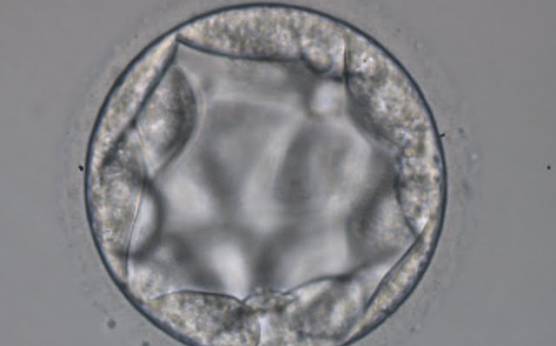
Figure 369
Blastocyst (Grade 3:3:2) with TE cells that in places are quite large and stretch over great distances to reach the next cell. No ICM can be identified.
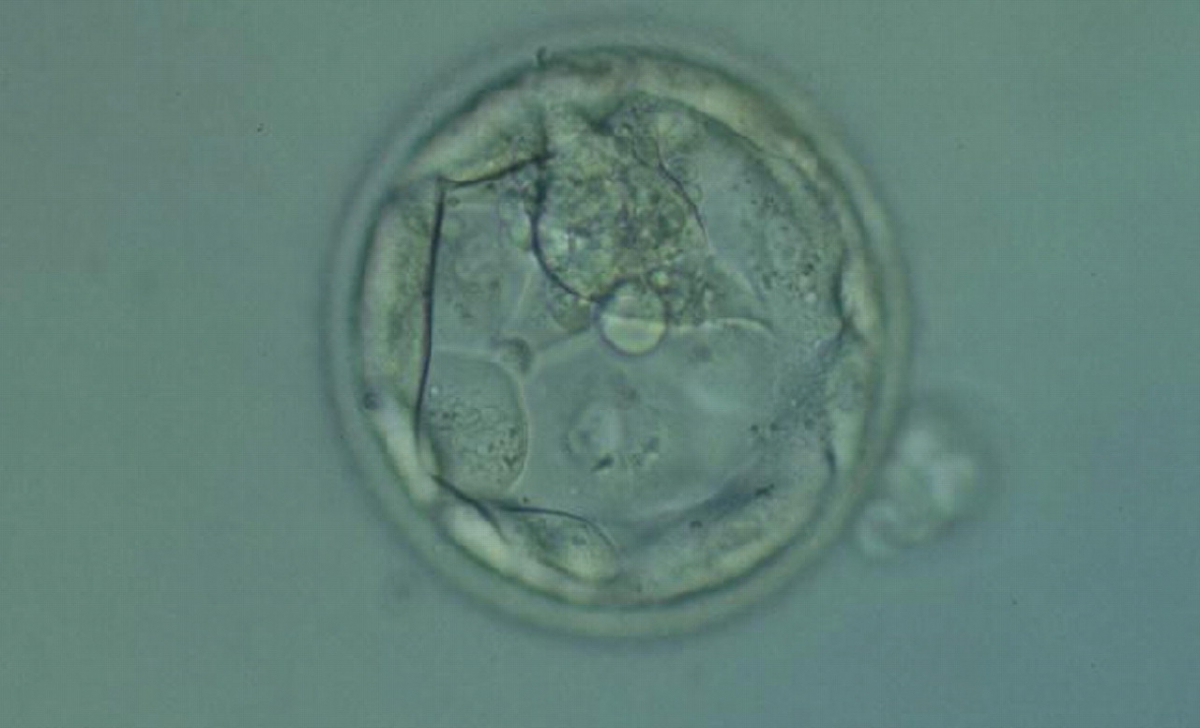
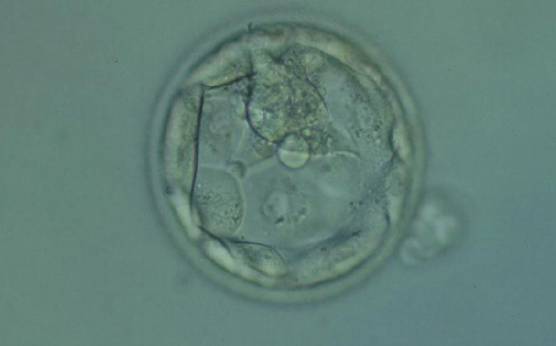
Figure 370
Hatching blastocyst (Grade 5.1.2) with few TE cells that form a loosely cohesive epithelium, and with a mushroom-shaped ICM at the 12 o'clock position in this view. The blastocyst was transferred but failed to implant.
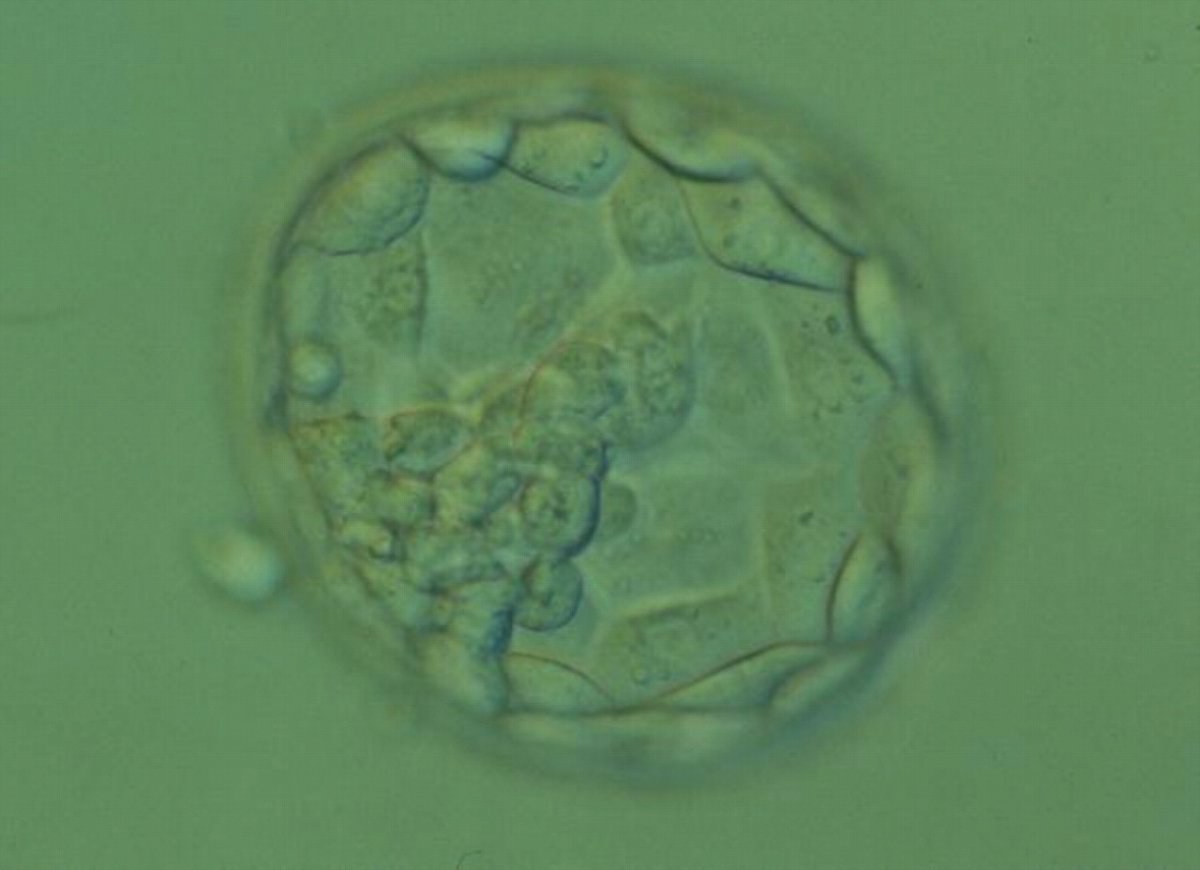
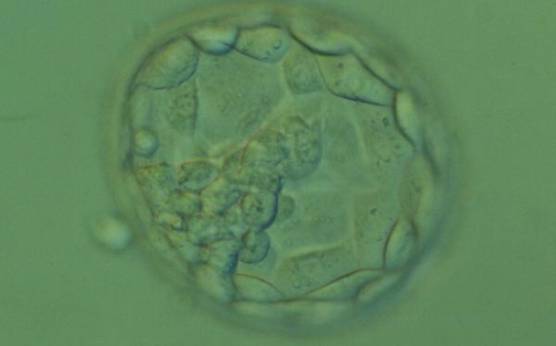
Figure 371
Hatching blastocyst (Grade 5.1.2). The TE cells vary in size with some cells quite large forming a loosely cohesive epithelium. A stellate ICM can be seen at the 8 o'clock position in this view. The blastocyst was transferred and failed to implant.
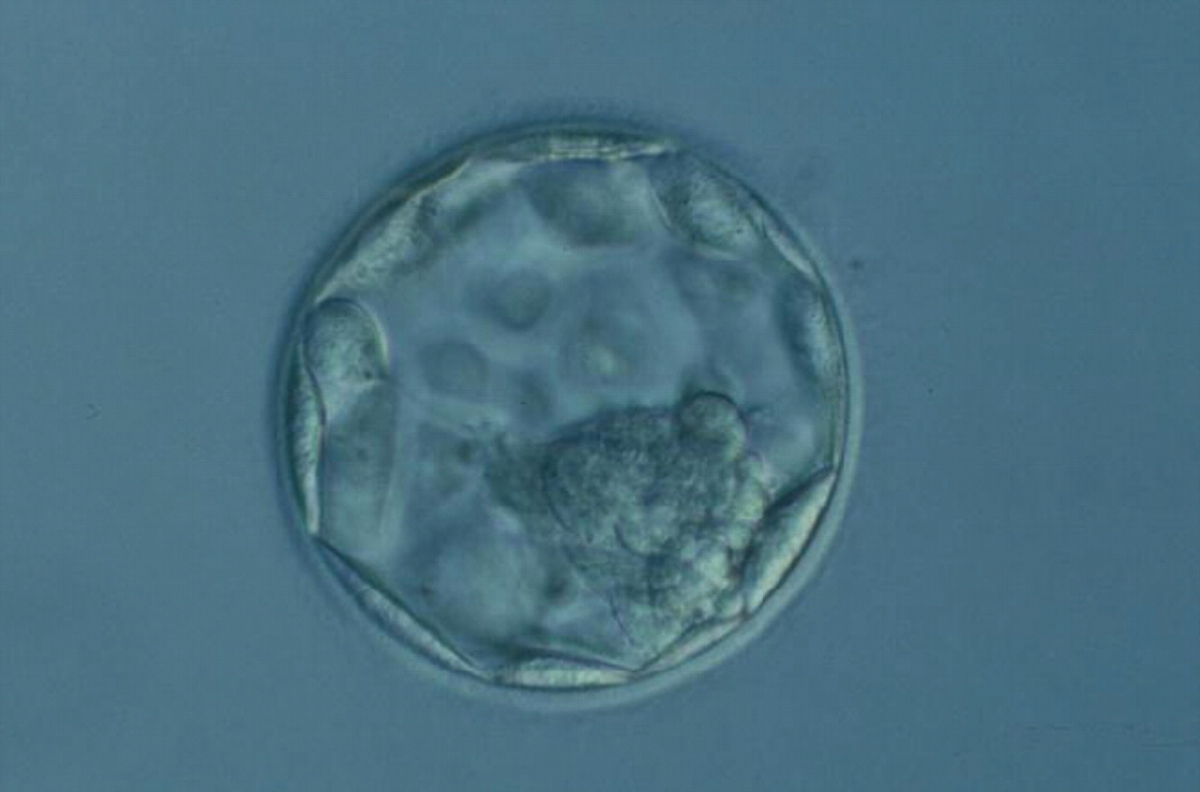
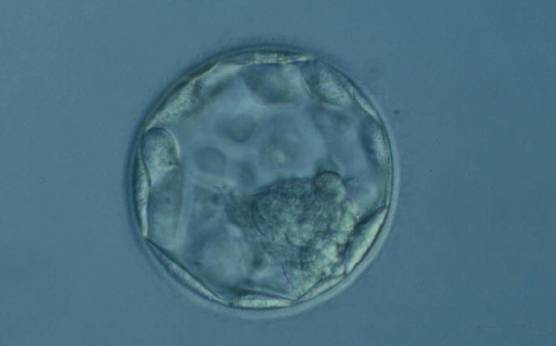
Figure 372
Expanded blastocyst (Grade 4:1:2). The TE cells vary in size with some cells quite large being particularly evident at the edge of the blastocyst where the cells stretch over some distance to reach their nearest neighbors. The blastocyst was transferred but the outcome is unknown.
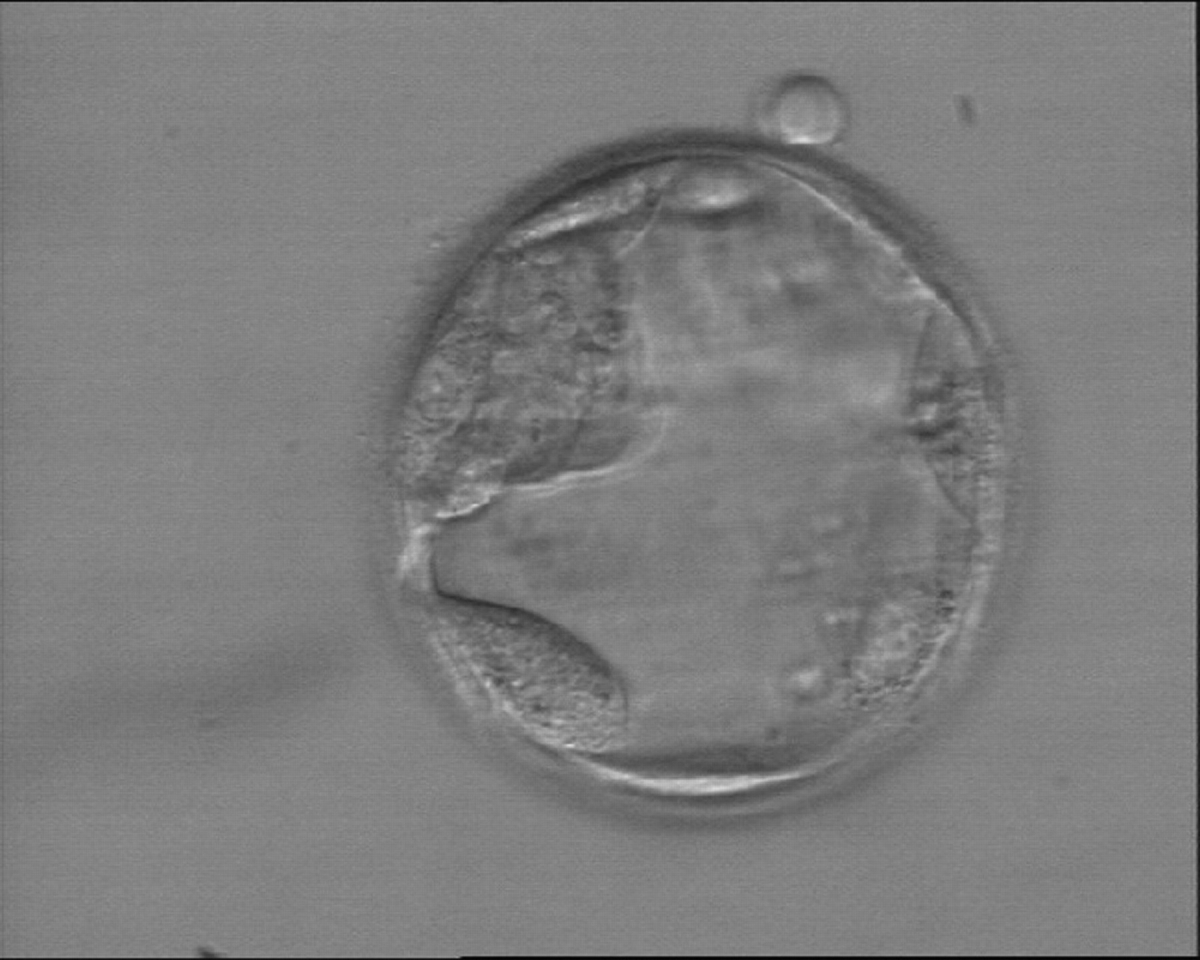
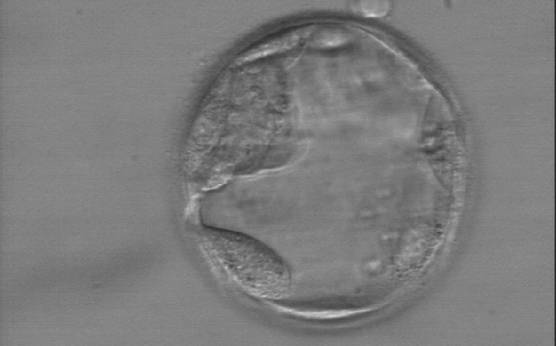
Figure 373
Early hatching blastocyst (Grade 5:1:3) with a very sparse TE that does not form a cohesive epithelium. The ICM is visible at the 10 o'clock position in this view.
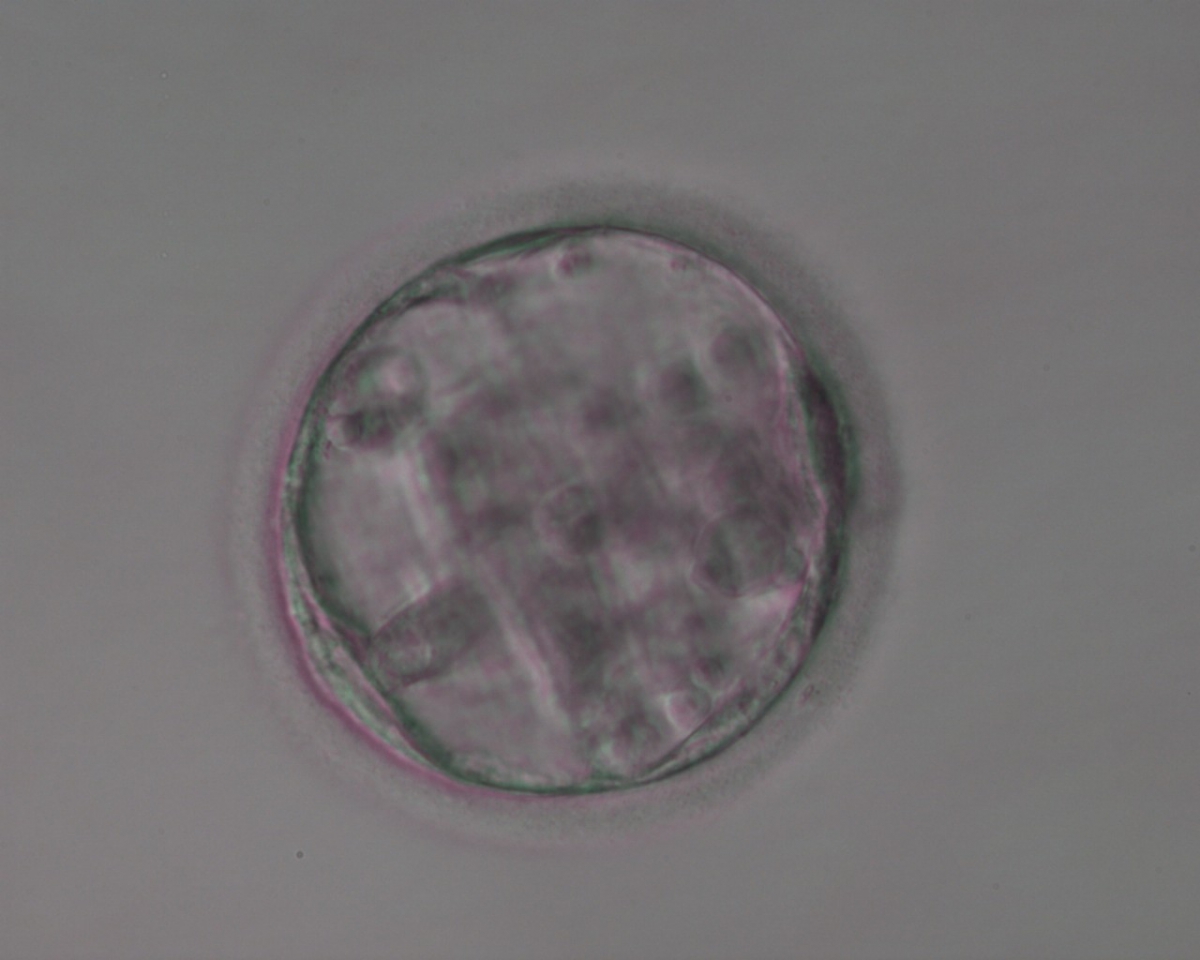
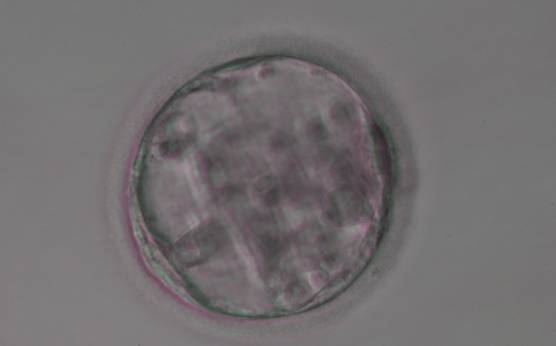
Figure 374
Blastocyst (Grade 3:3:3) with sparse TE that does not form a cohesive epithelium. The ICM is not clearly identifiable.
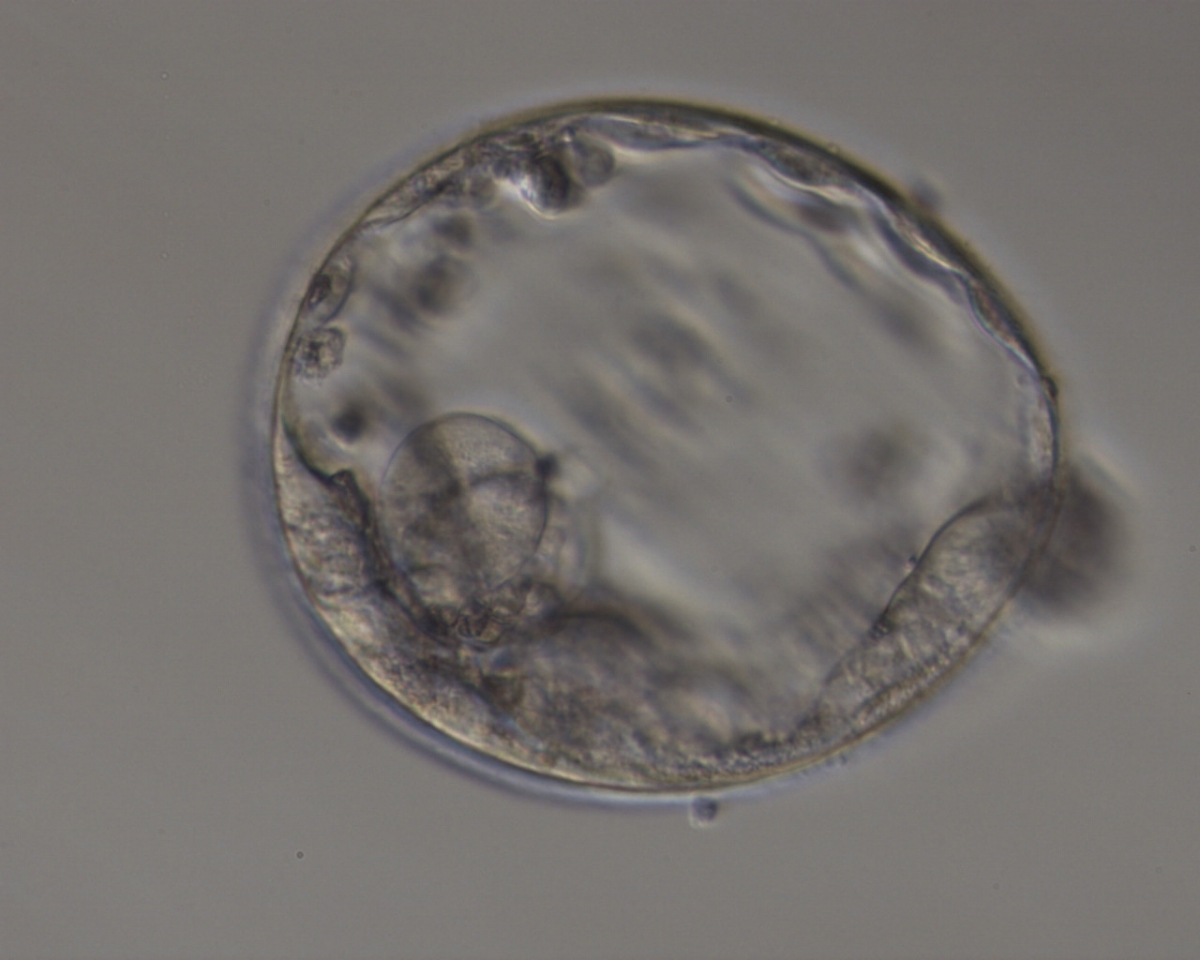
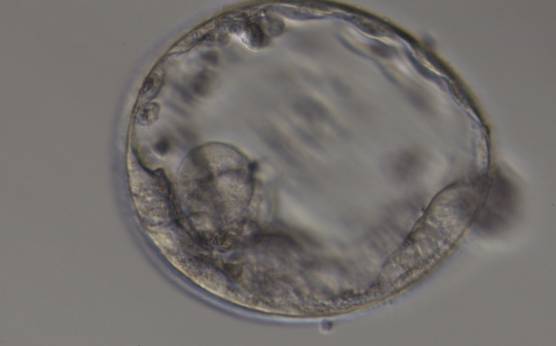
Figure 375
Expanded blastocyst (Grade 4:3:3) with sparse TE that does not form a cohesive epithelium. The ICM is hardly distinguishable despite the expansion of the blastocoel cavity.

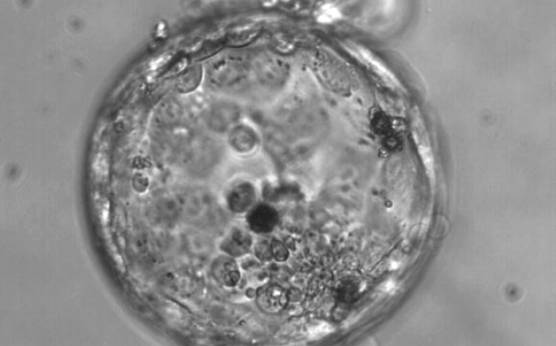
Figure 376
Hatching blastocyst (Grade 5:3:3). The TE varies in size and does not form a cohesive epithelium. Several loosely cohesive ICM cells can be seen at the 5 o'clock position in this view. There are several dark degenerate foci within the blastocyst.
Fong et al. (2001) reported the detailed ultrastructural appearance of TE cells from naturally hatched and enzymatically hatched blastocysts. There was no difference in the ultrastructural appearance of the TE cells following exposure to enzyme: outer tight and adherent junctions, desmosomes, gap junctions, microvilli on the free surfaces, oval-to-tubular mitochondria with well-developed cristae typical of blastocysts, rough endoplasmic reticulum, Golgi complexes, occasional centrioles associated with microtubules, lysosomes, multivesicular bodies and spherical lipid globules present in many cells. Dark granules can be observed in many TE cells at the light microscopic level which are likely to be the lipid globules (Figs 377–379).
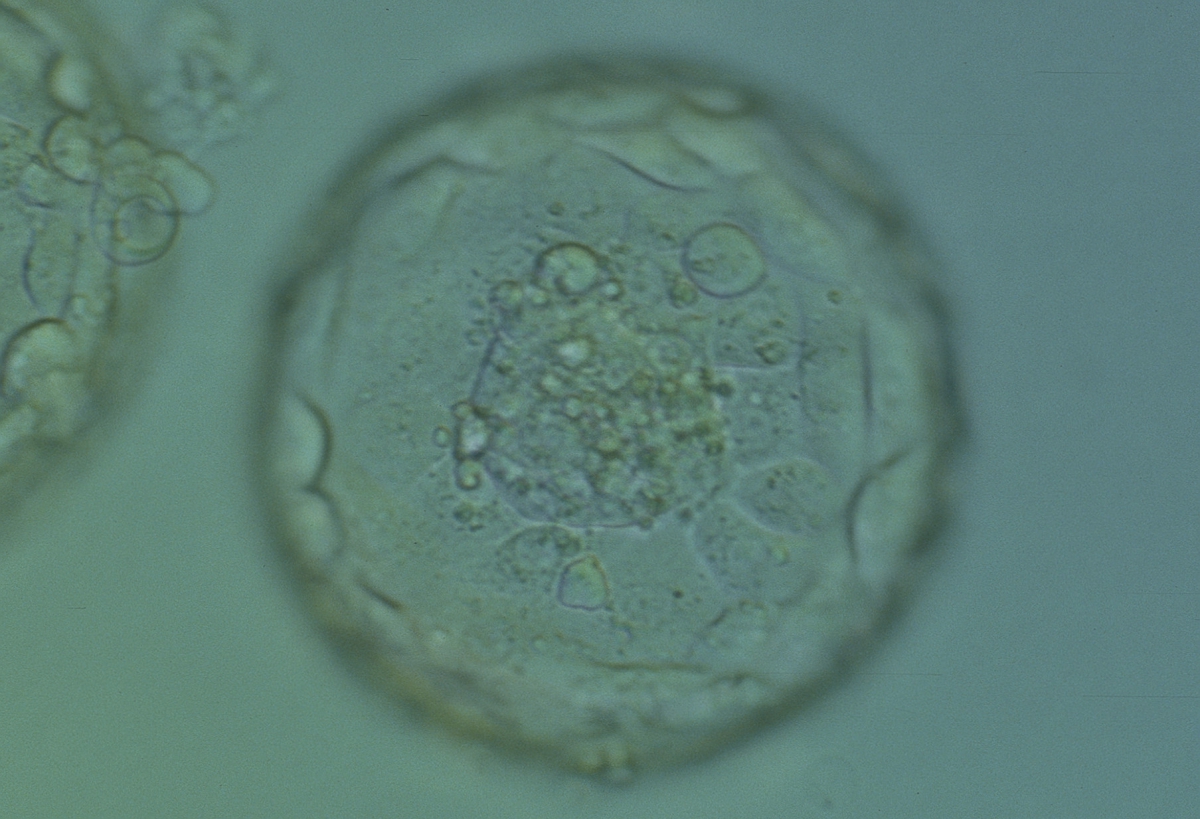
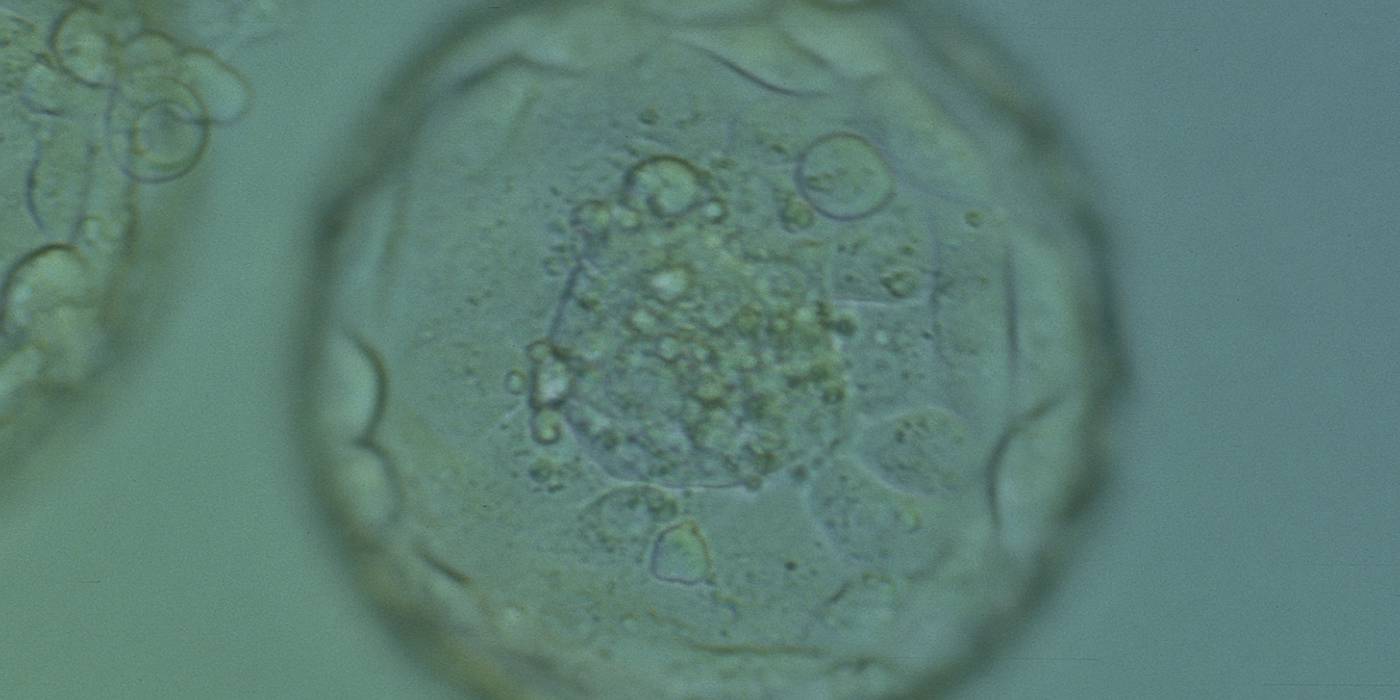
Figure 377
Expanded blastocyst (Grade 4:1:1). Note several dark granules within the majority of the TE cells. The ICM has several fragments. The blastocyst was transferred but failed to implant.
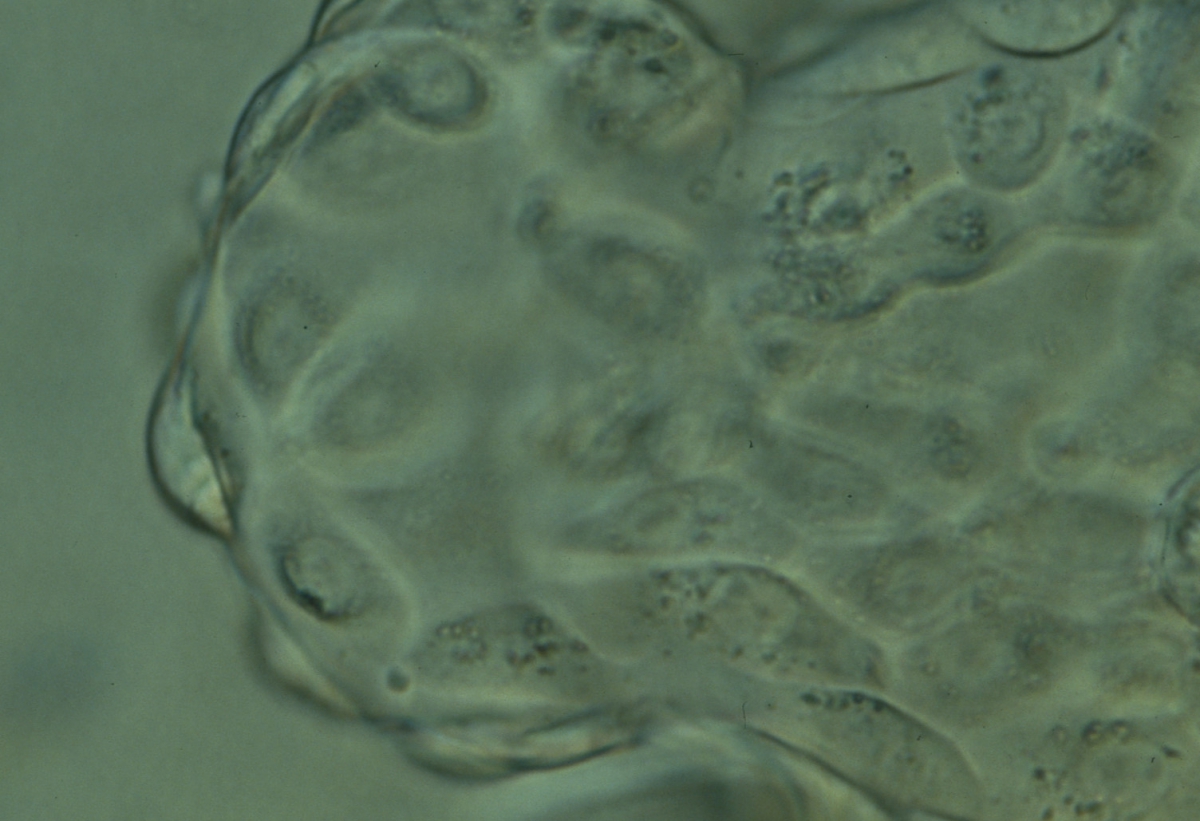
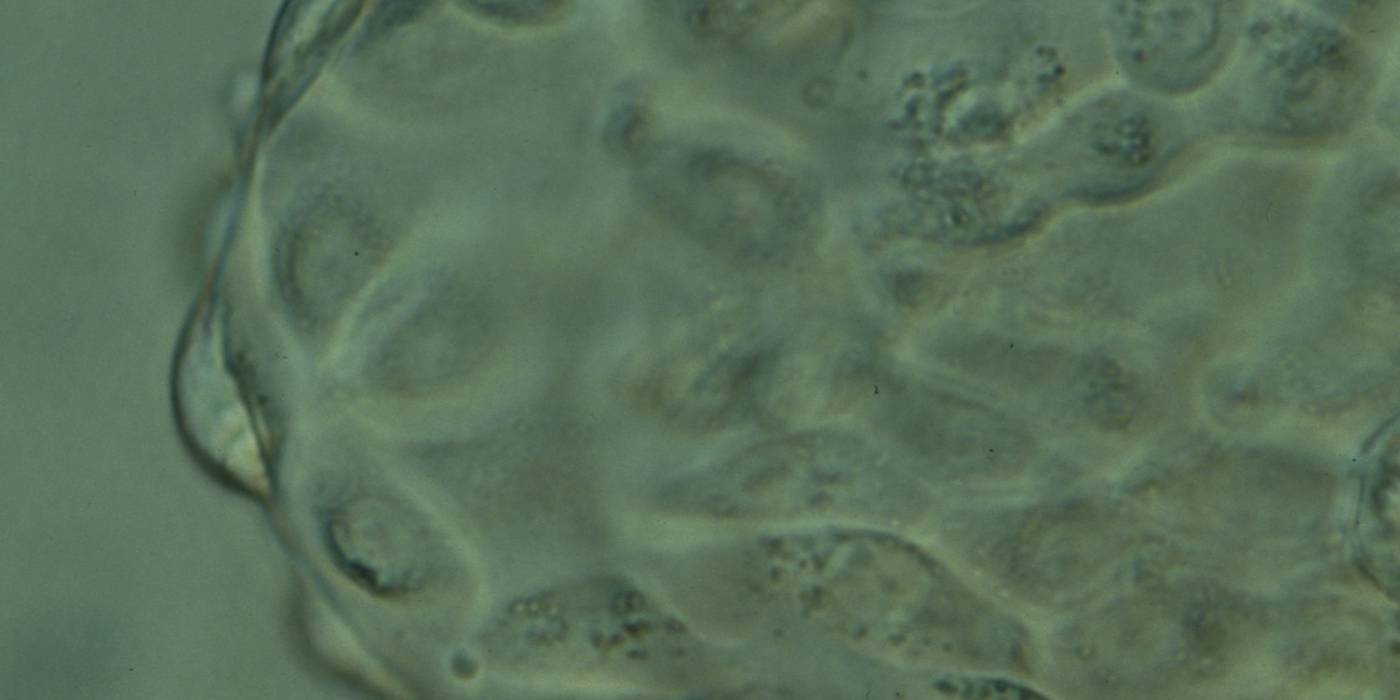
Figure 378
Hatching blastocyst (Grade 5:1:1) showing the herniating TE cells at high magnification. Many of the TE cells contain several dark granules. The blastocyst was transferred but the outcome is unknown.
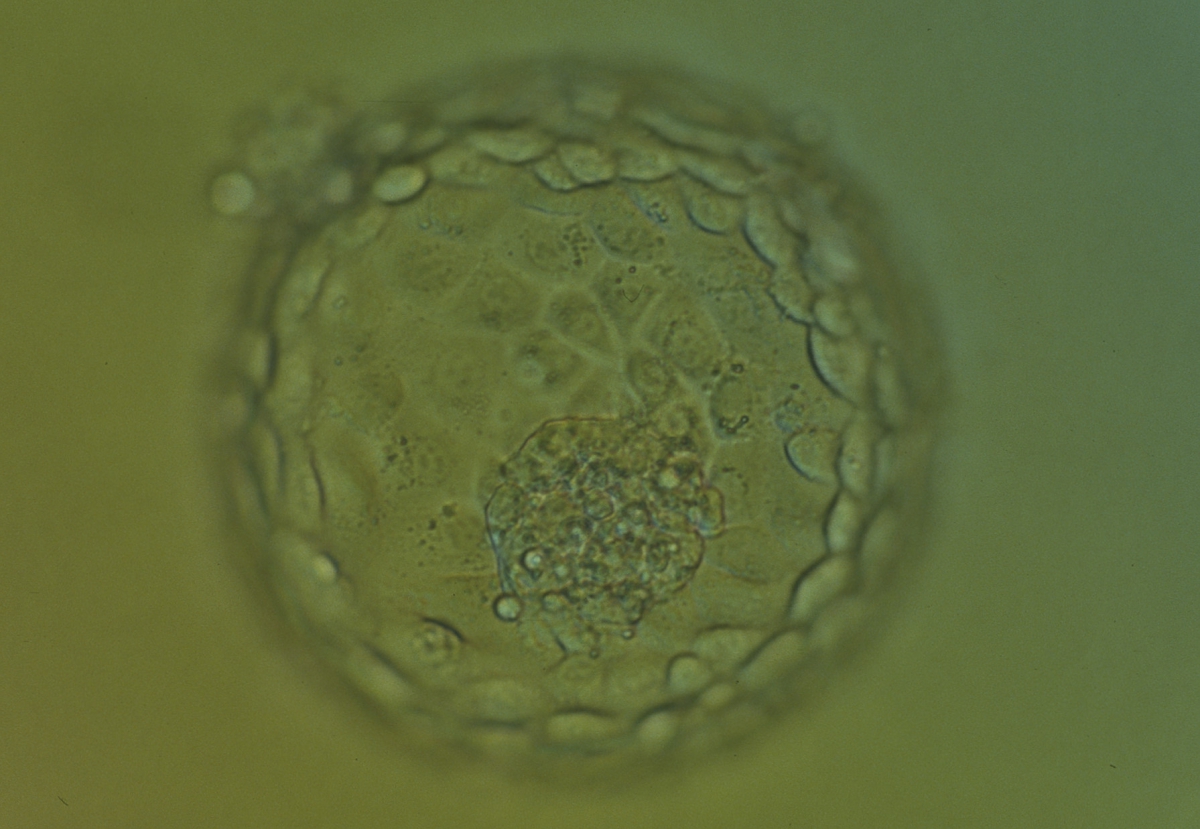
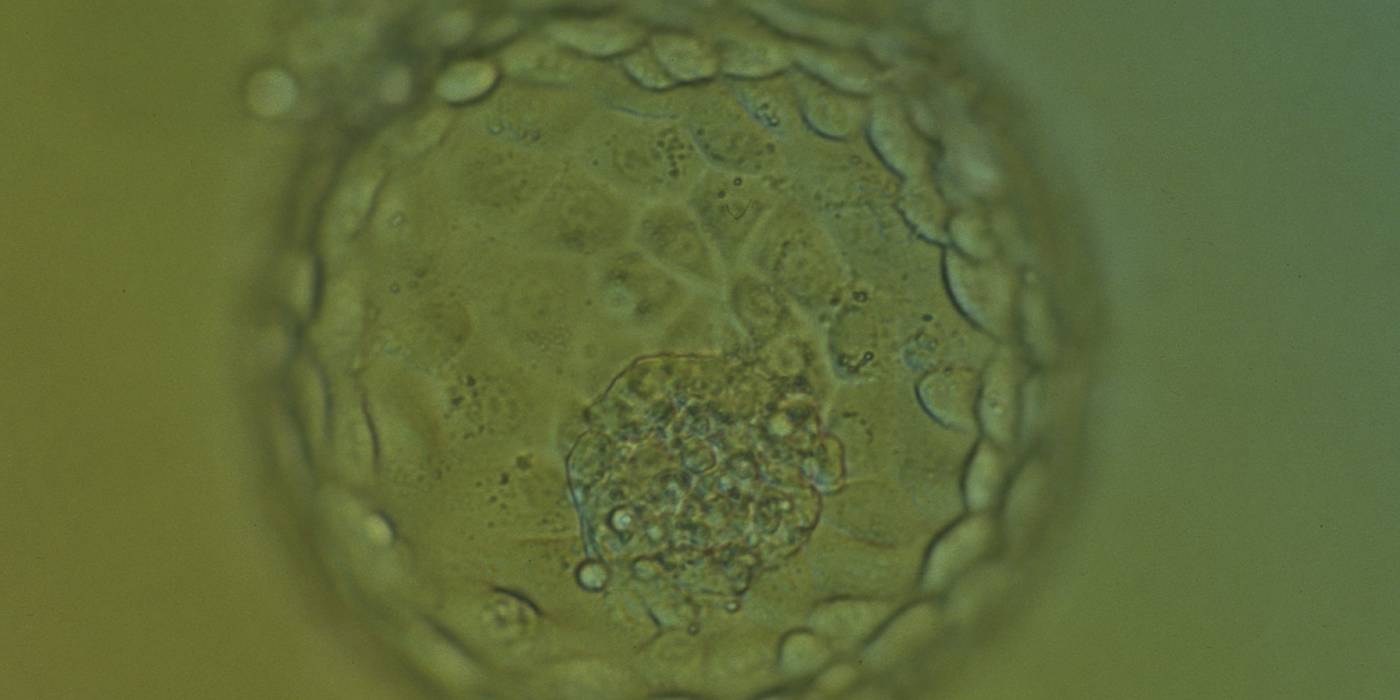
Figure 379
Hatching blastocyst (Grade 5:1:1) showing many of the TE cells contain dark granules. Several fragments are associated with a centrally positioned, compact ICM. The blastocyst was transferred and implanted.
Article references:
Ahlström A, Westin C, Reismer E, Wikland M, Hardarson T. Trophectoderm morphology: an important parameter for predicting pregnancy and birth after single blastocyst transfer. Hum Reprod 2011;26:3289-3296.
Abstract/FREE Full Text
Aplin JD. The cell biological basis of implantation. Baillieres Best Pract Res Clin Obstet Gynaecol 2000;14:757-764.
CrossRef | Medline | Google Scholar
Fong CY, Bongso A, Sathananthan H, Ho J, Ng SC. Ultrastructural observations of enzymatically treated human blastocysts: zona-free blastocyst transfer and rescue of blastocysts with hatching difficulties. Hum Reprod 2001;16:540-546.
Abstract/FREE Full Text
Richter KS, Harris DC, Daneshmand ST, Shapiro BS. Quantitative grading of a human blastocyst: optimal inner cell mass size and shape. Fertil Steril 2001;76:1157-1167.
CrossRef | Medline | Web of Science | Google Scholar
Sathananthan H, Menezes J, Gunasheela S. Mechanics of human blastocyst hatching in vitro. Reprod BioMed Online 2003;7:228-234.
CrossRef | Medline | Google Scholar
Zaninovic N, Berrios R, Clarke RN, Bodine R, Ye Z, Veeck LL. Blastocyst expansion, inner cell mass (ICM) formation, and trophectoderm (TM) quality: is one more important for implantation? Fertil Steril 2001;76:S8.
Google Scholar
