D. Nucleolar precursor bodies
D.1 NPBs: aligned/scattered/differential alignment between PNs
As PNs form after fertilization, there is polarized distribution in the chromatin into the furrow between them (Van Blerkom et al., 1997). As NPBs are attached to the chromatin, they should also polarize or align with it implying that, if there is correct chromatin polarization, the NPBs will appear polarized as well (Figs 151–154). Due to the dynamics of this event, some zygotes show symmetry in appearance of the PNs, but a delay in the alignment of the chromatin into the furrow, or onto the mitotic plate (Figs 155–157). NPB and PN progression can occur even in highly dysmorphic zygotes (Fig. 158).
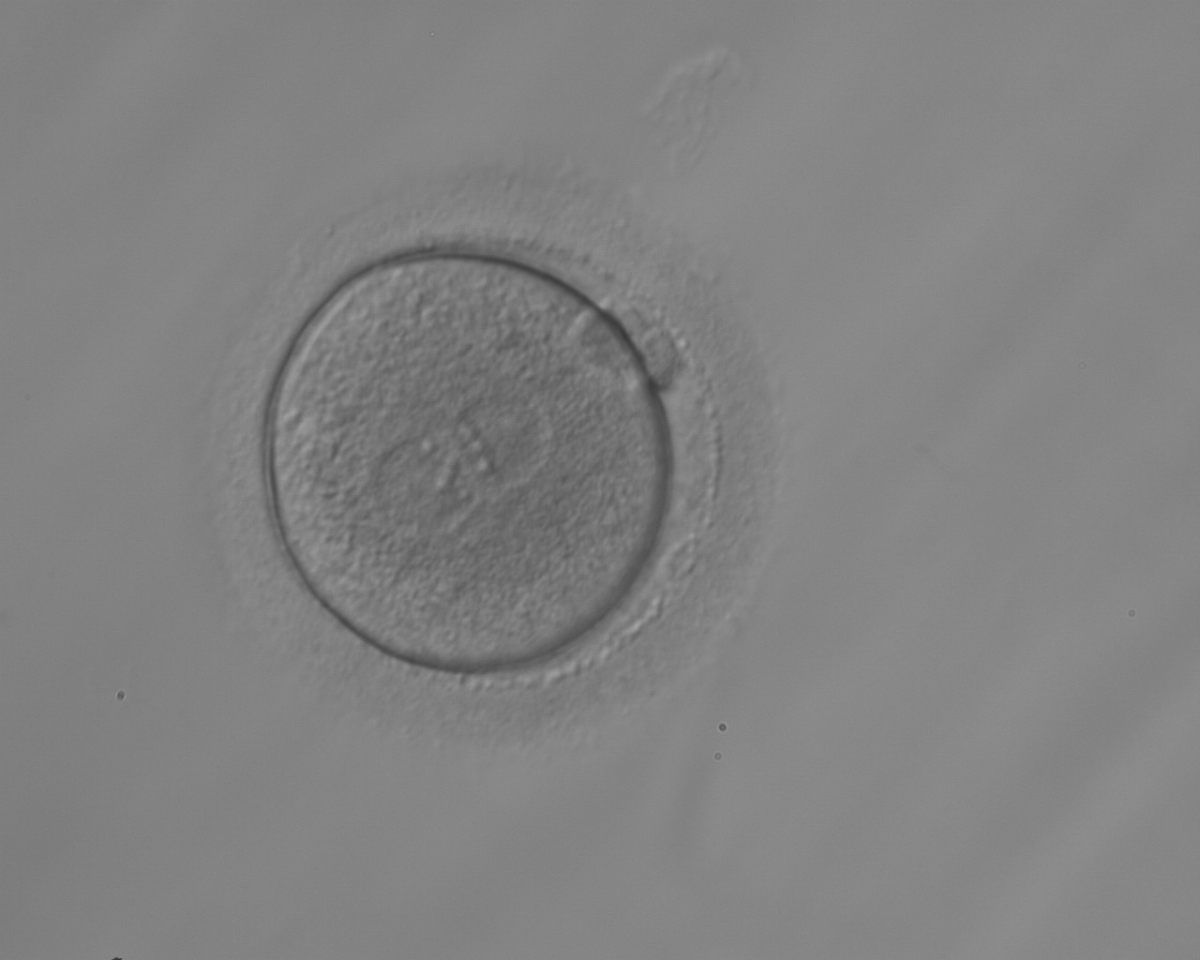
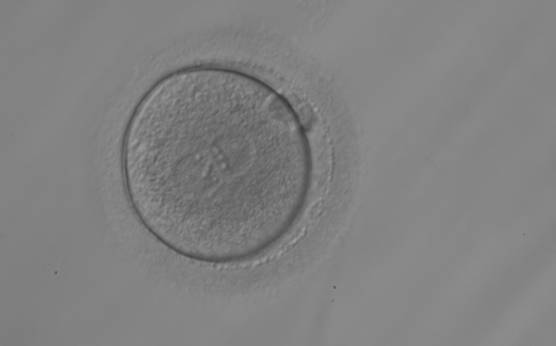
Figure 151
A zygote observed 18 h post-ICSI with equal numbers of NPBs aligned at the PN junction (200× magnification). It was transferred but the outcome is unknown.
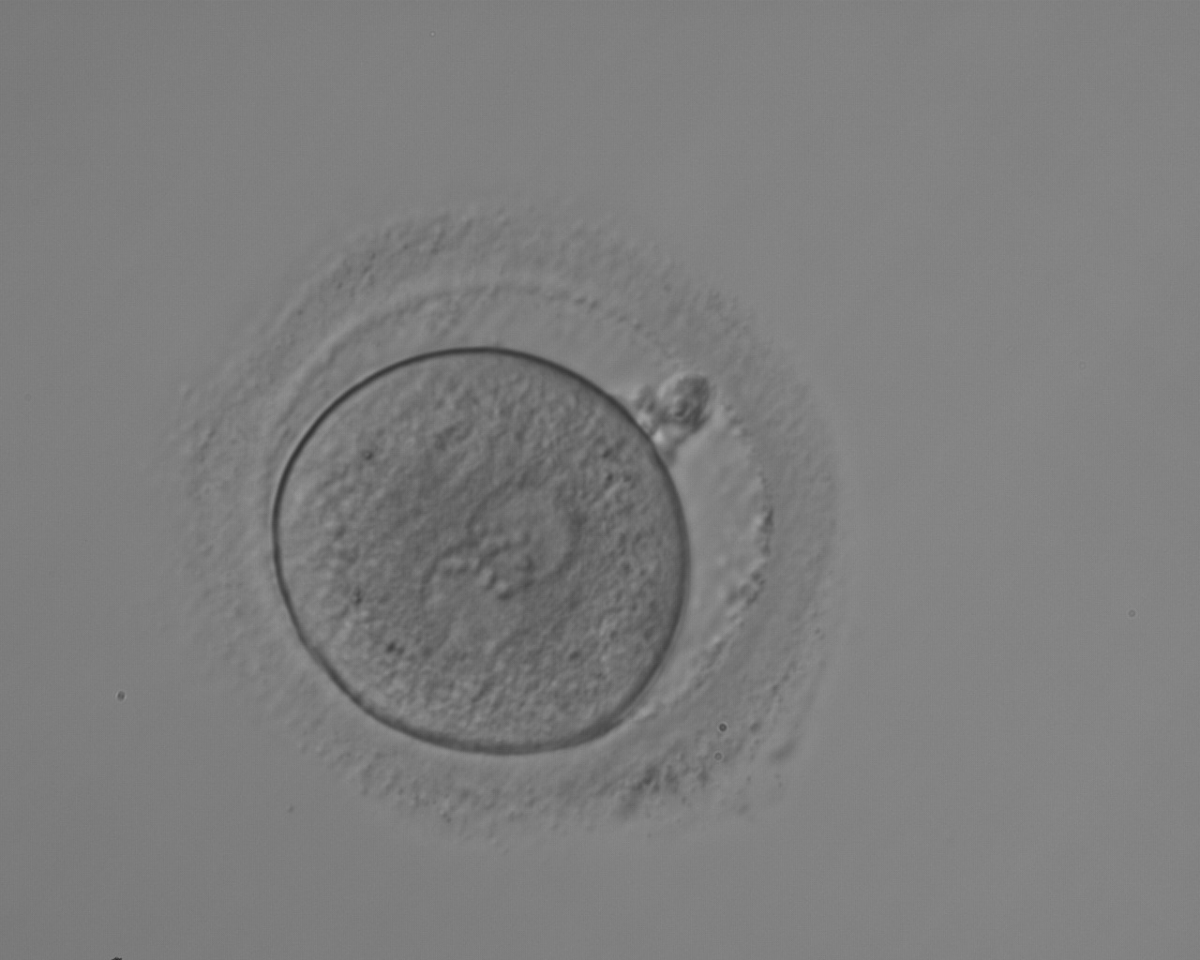
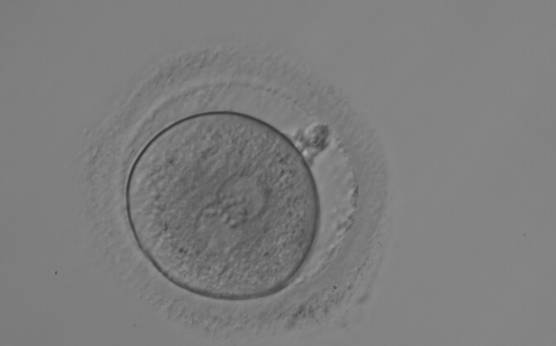
Figure 152
A zygote observed 18 h post-ICSI with equal numbers of NPBs aligned at the PN junction (200× magnification). It shows a large PVS and an irregular ZP. The polar bodies are fragmented and overlapping in this view. It was transferred but the outcome is unknown.
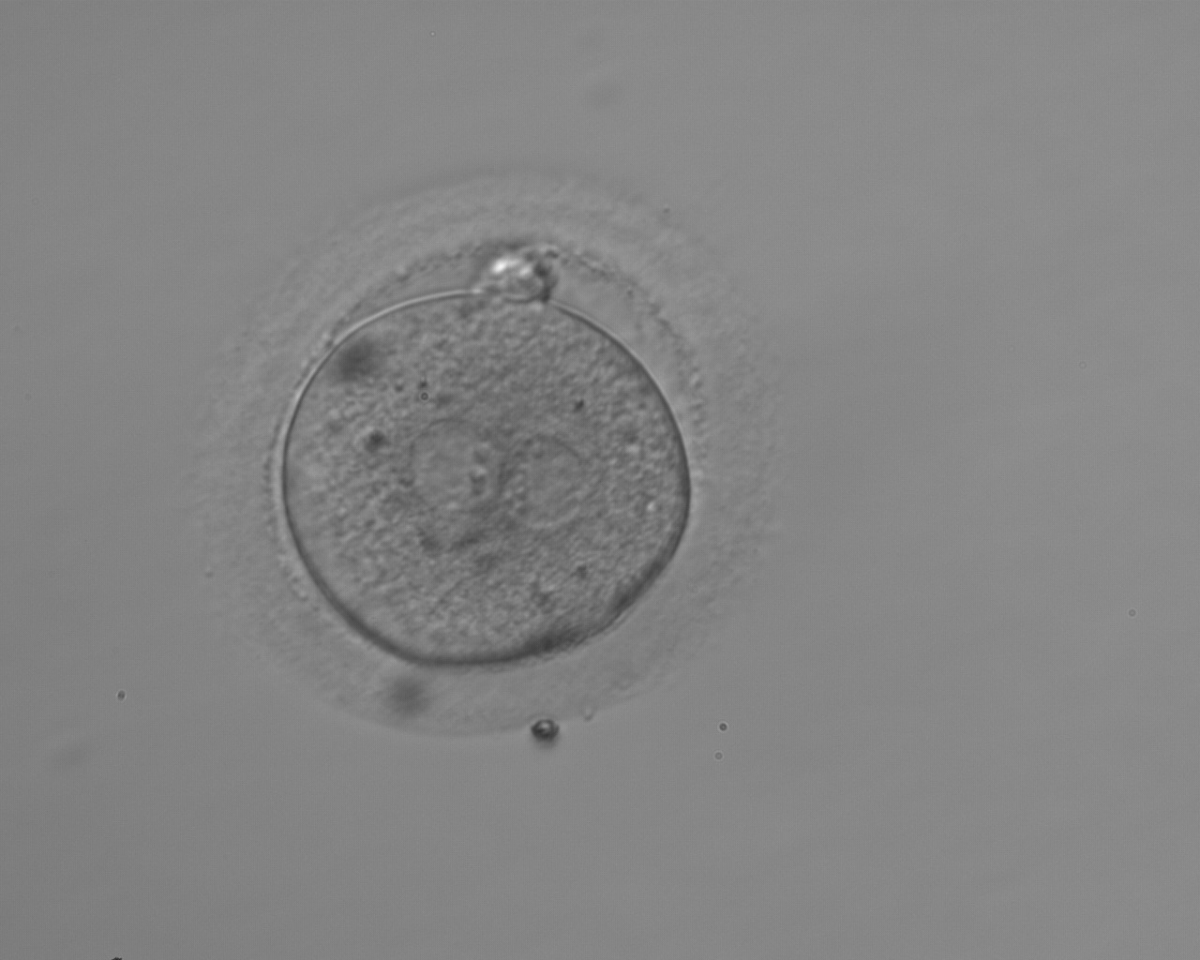
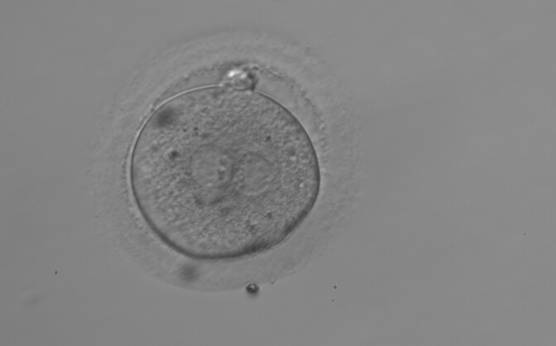
Figure 153
A zygote observed 18 h post-ICSI with NPBs aligned at the PN junction (200× magnification). NPBs differ in number and size between PNs. The polar bodies are overlapped in this view. It was discarded due to subsequent abnormal cleavage.
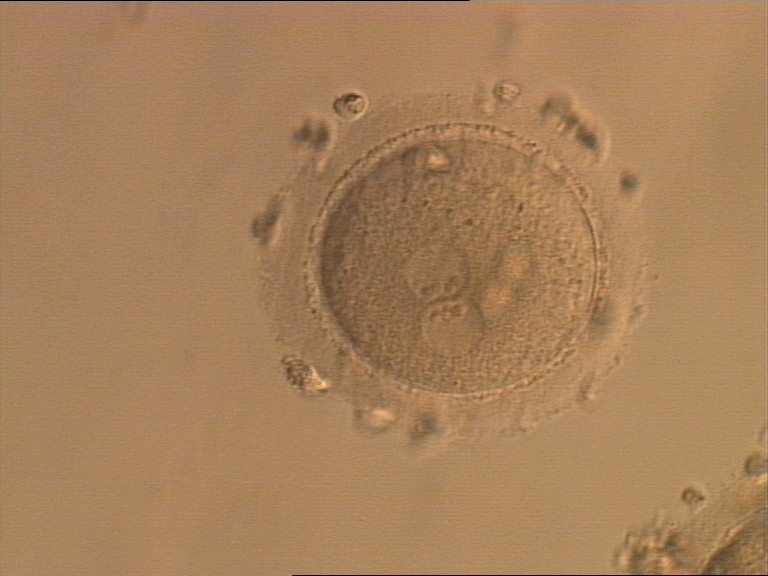
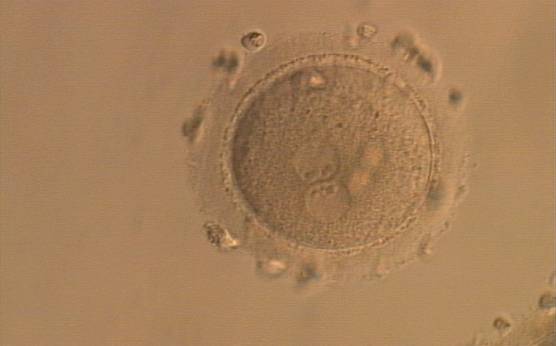
Figure 154
A zygote generated by ICSI, displaying two perfectly juxtaposed PNs within which the NPBs are aligned, have the same size and are similar in number (400× magnification). It was transferred but the outcome is unknown.
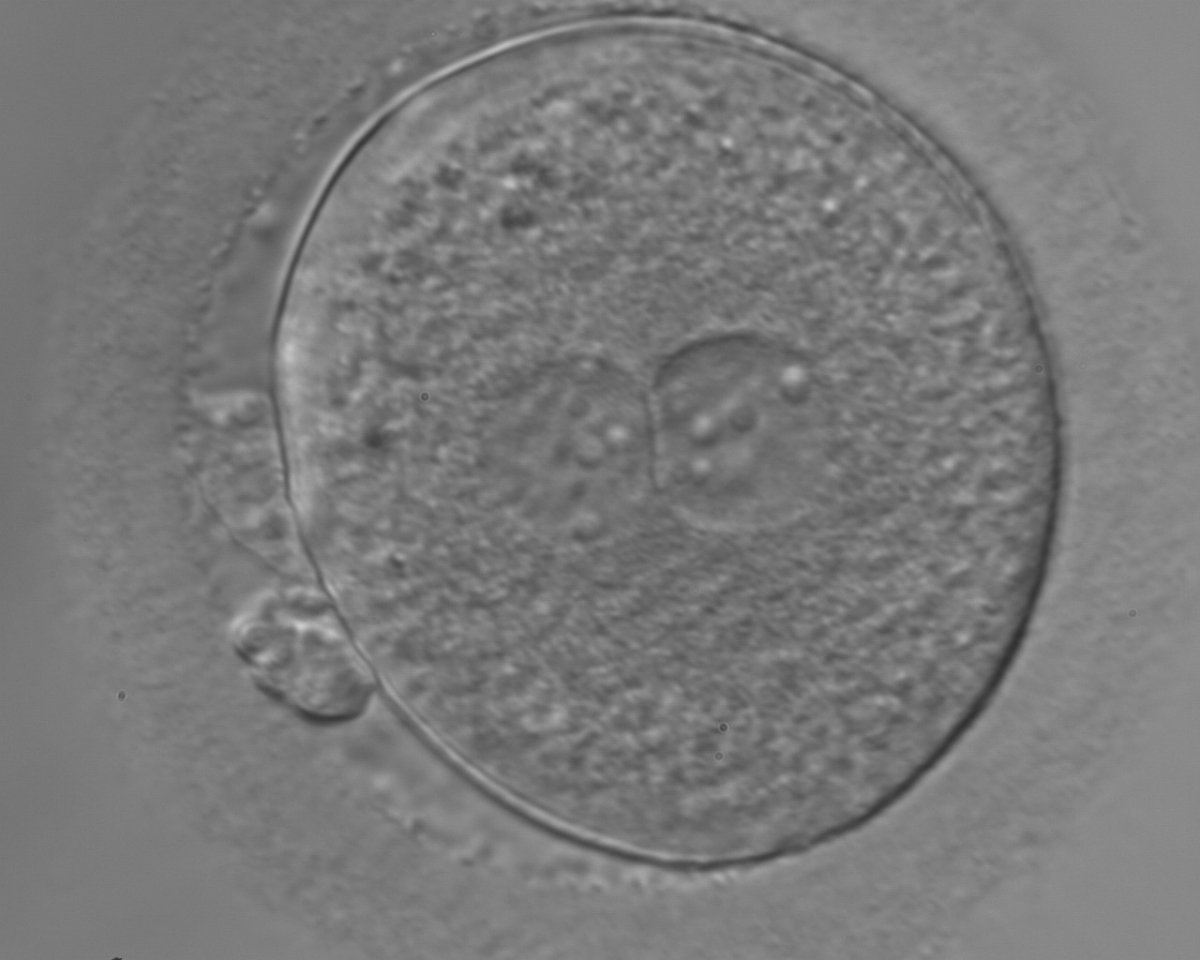
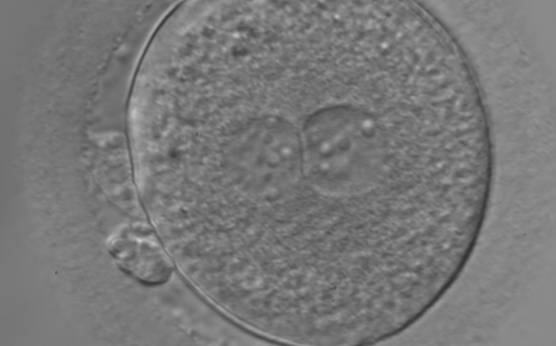
Figure 155
An ICSI zygote with large-sized NPBs scattered with respect to the PN junction (400× magnification). The cytoplasm appears granular. It was transferred but failed to implant.
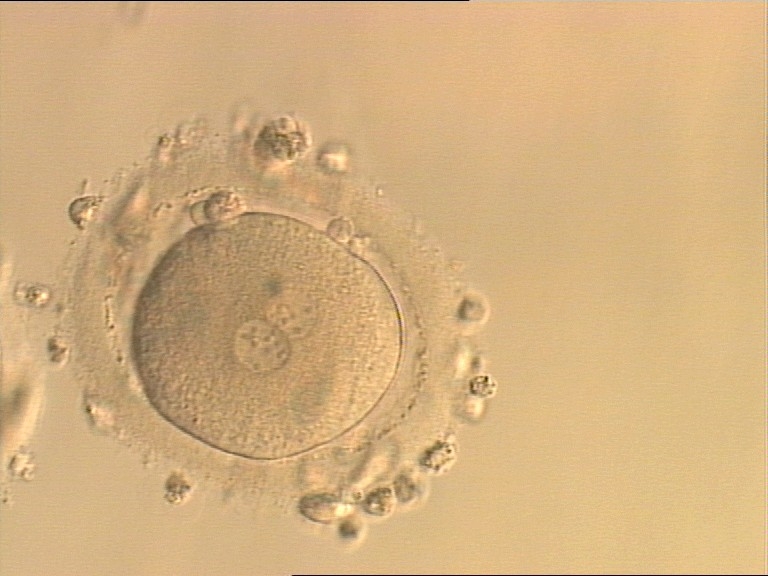
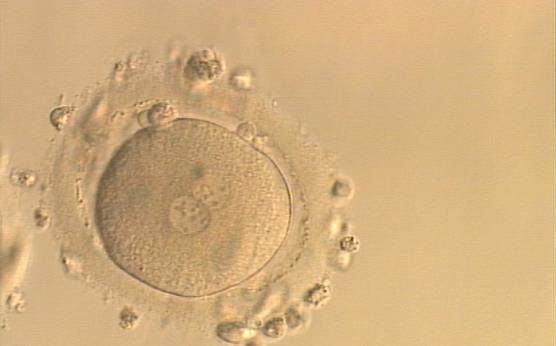
Figure 156
An ICSI zygote with NPBs scattered with respect to the PN junction (400× magnification). Polar bodies are fragmented; there is debris in the PVS. It was transferred but implantation outcome is unknown.
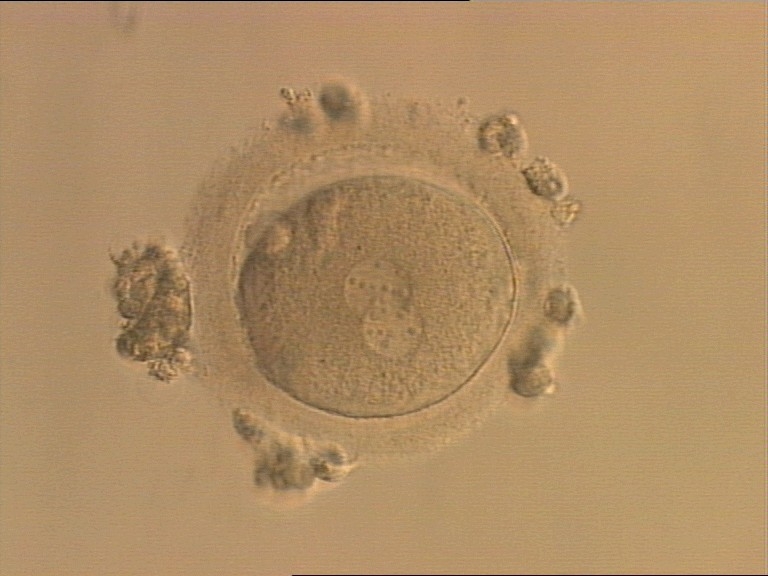
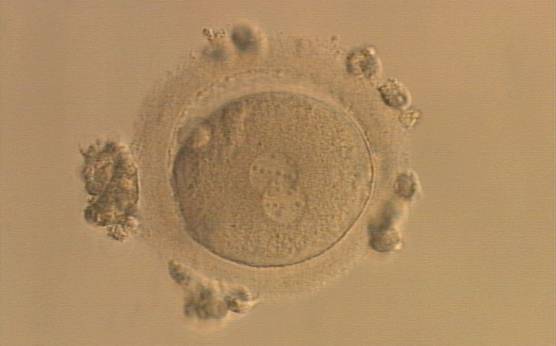
Figure 157
An ICSI zygote displaying partially overlapping PNs in this view. NPBs are scattered in both PNs (400× magnification). It was transferred but the outcome is unknown.
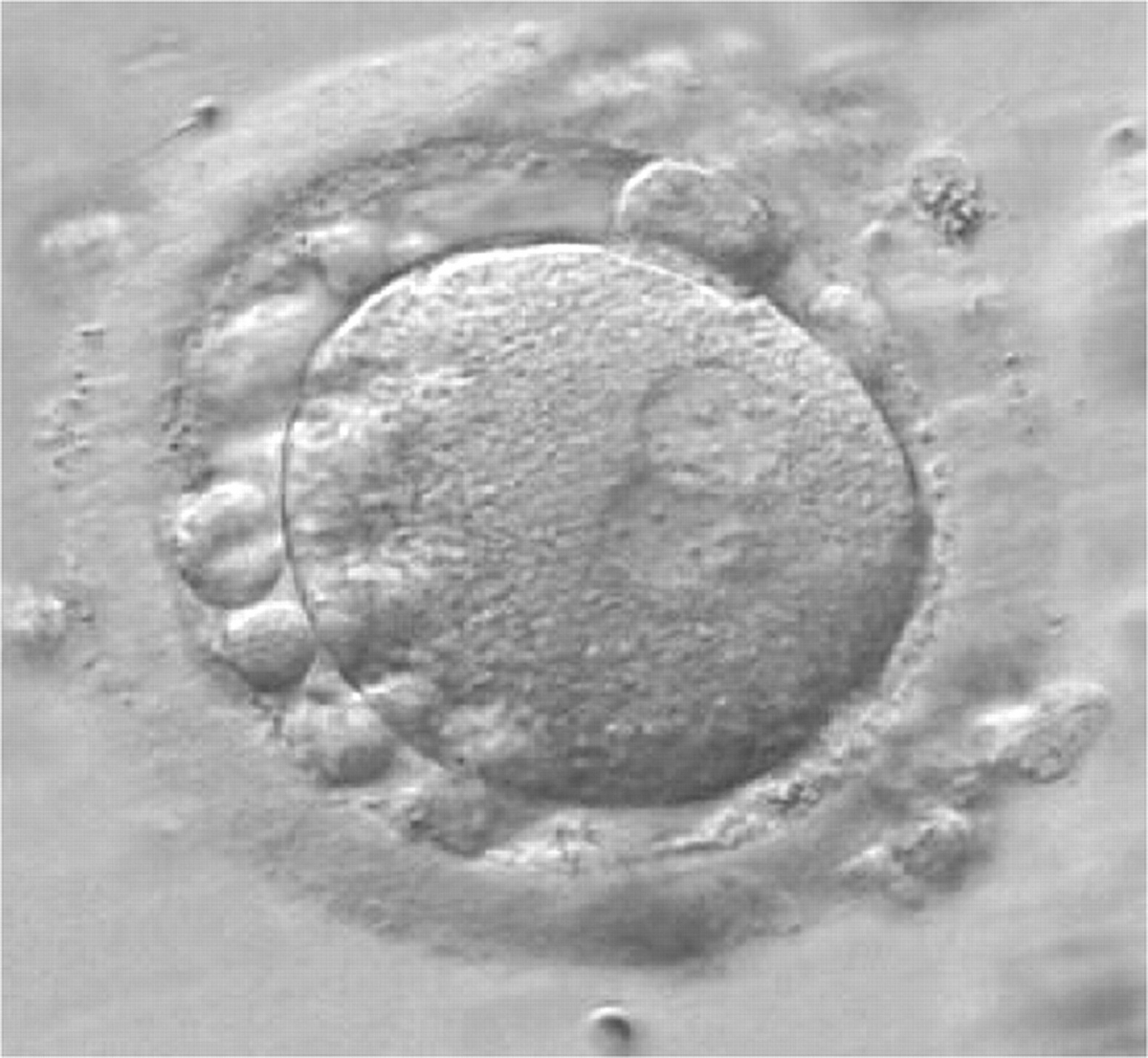
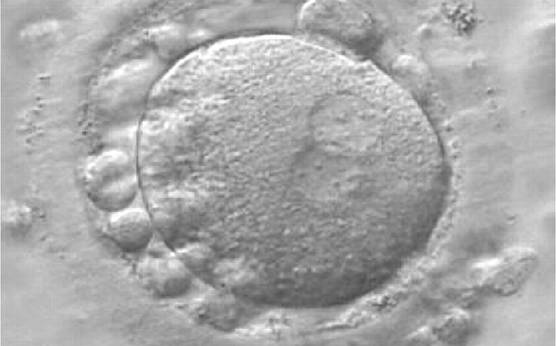
Figure 158
A zygote generated by IVF at 16.40 h post-insemination (400× magnification). It shows small-sized NPBs scattered with respect to the PN junction and a lot of large cellular debris in the PVS. It was discarded.
During the progression of the cell cycle, NPBs change in number, size and distribution (Fig. 101) and finally they disappear shortly before syngamy and initiation of the first cleavage division (Fig. 149). Recent investigations by time-lapse imaging have shown that NPBs are highly dynamic and that a characteristic NPB pattern may change within a short period of time (Montag et al., 2011).
The potential use of the arrangement of NPBs in both PNs regarding size, number and symmetry was initially investigated as a major part of PN scoring in the late 90s (Scott and Smith, 1998). Several publications found a benefit of PN scoring and especially the distribution of NPBs with the outcome of assisted reproduction treatment (Tesarik and Greco, 1999; Scott et al., 2000; Balaban et al., 2001; Montag and van der Ven, 2001; Ebner et al., 2003; Senn et al., 2006). Others have reported no benefit (Payne et al., 2005; James et al., 2006; Nicoli et al., 2007; Brezinova et al., 2009).
NPBs' morphology is influenced by the patient's age and there is also a temporal difference in morphology between zygotes from IVF compared with ICSI. Therefore, comparative PN/NPB scoring should be performed according to strict guidelines considering timing, patient characteristics and insemination technique employed.
Many cell cycle control proteins are located in the nucleolus, and it has been shown in mitotic cells that asymmetry in number and pattern of NPBs is associated with abnormal cell cycles and ultimately with abnormal development (Pedersen, 1998). It is plausible that asymmetry between PNs in zygotes (Figs 159–162) can lead to abnormal development with an increase in fragmentation and abnormal cleavage, and reduced viability (Scott, 2003). Nevertheless, implantation can occur (Figs 163 and 164).
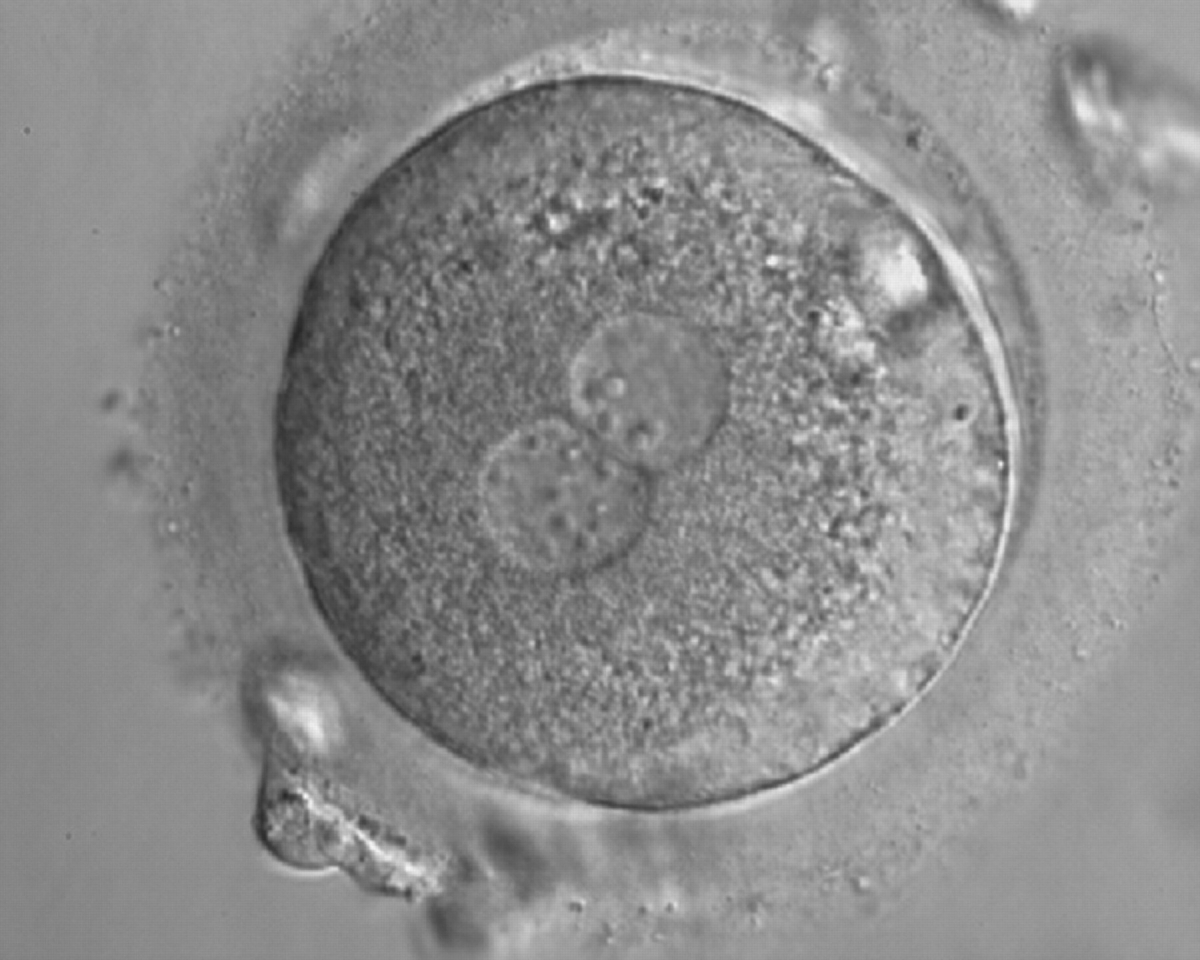
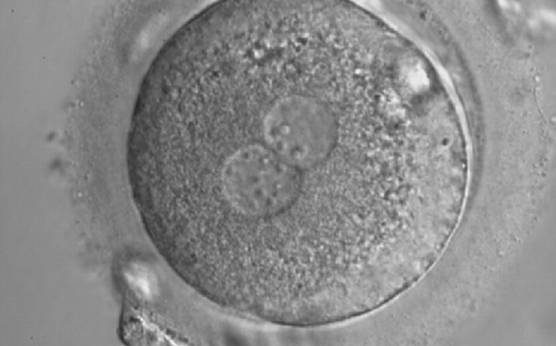
Figure 159
A zygote with inequality in numbers and alignment of NPBs. It was cryopreserved.
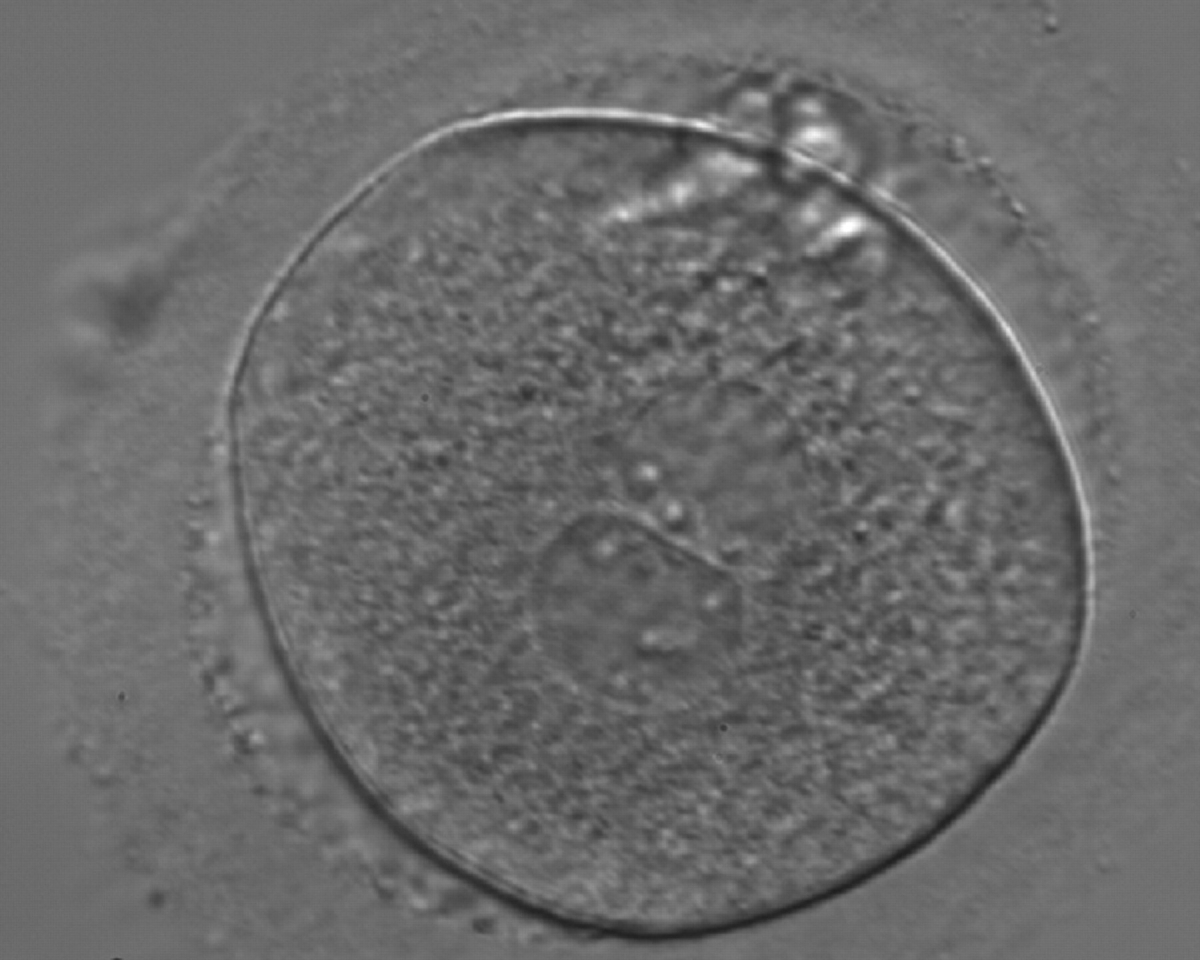
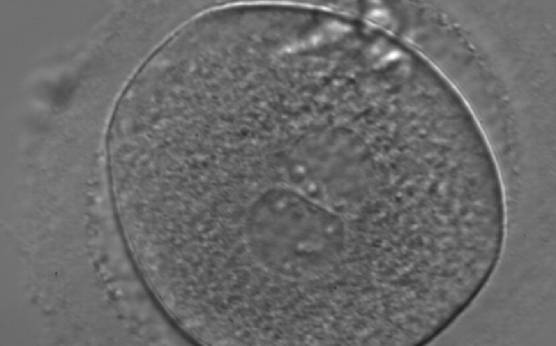
Figure 160
A zygote generated by ICSI with inequality in numbers and alignment of NPBs (400× magnification). NPBs are aligned in one PN and scattered in the other. Due to poor development, it was discarded.
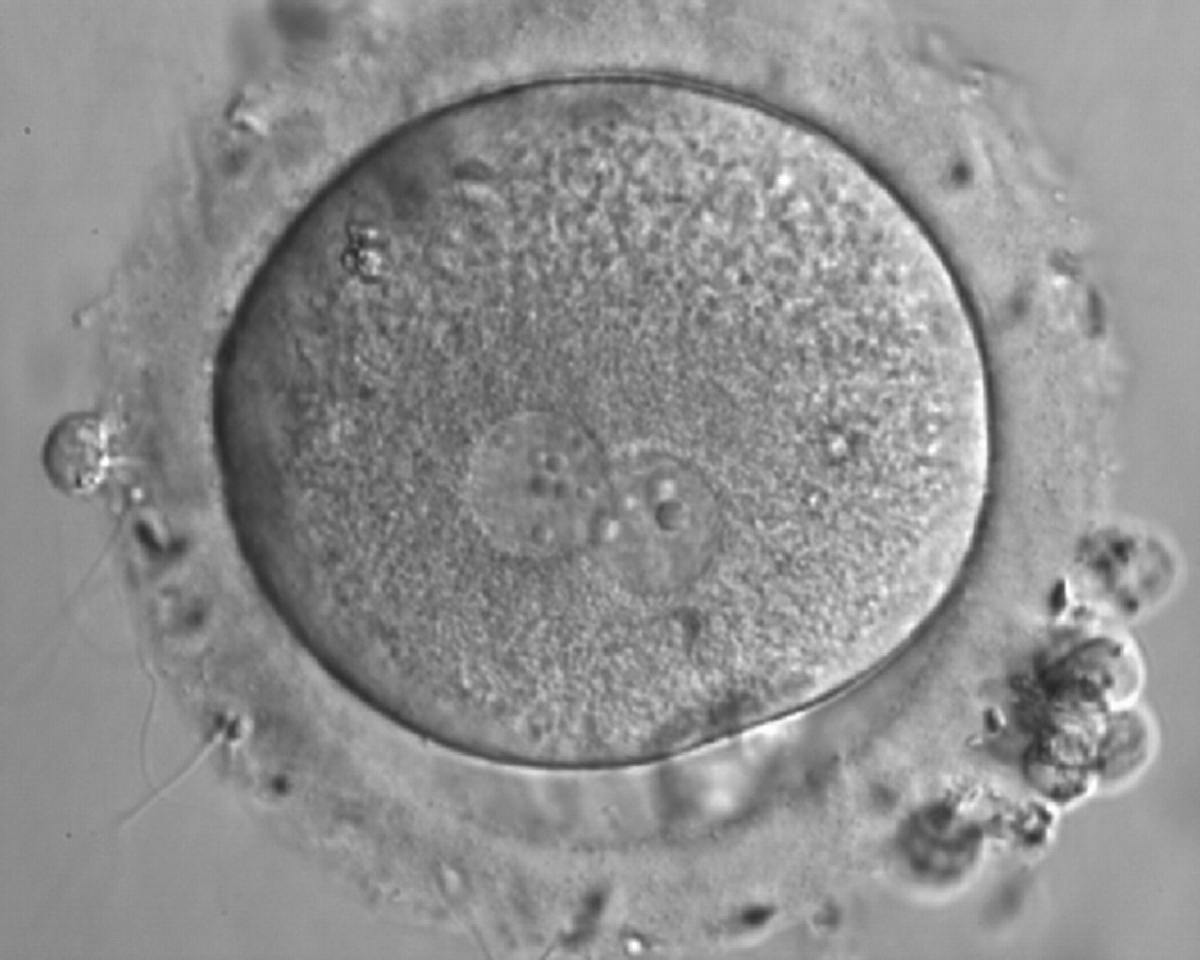
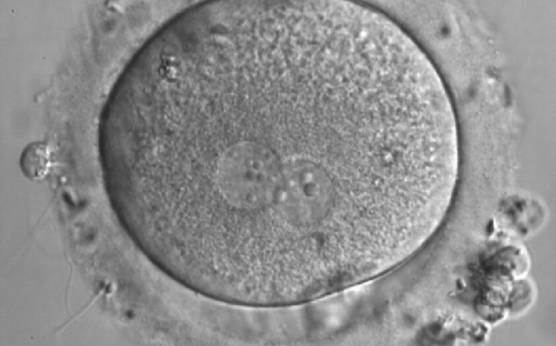
Figure 161
A zygote generated by IVF with inequality in numbers and alignment of NPBs (600× magnification). It was discarded.
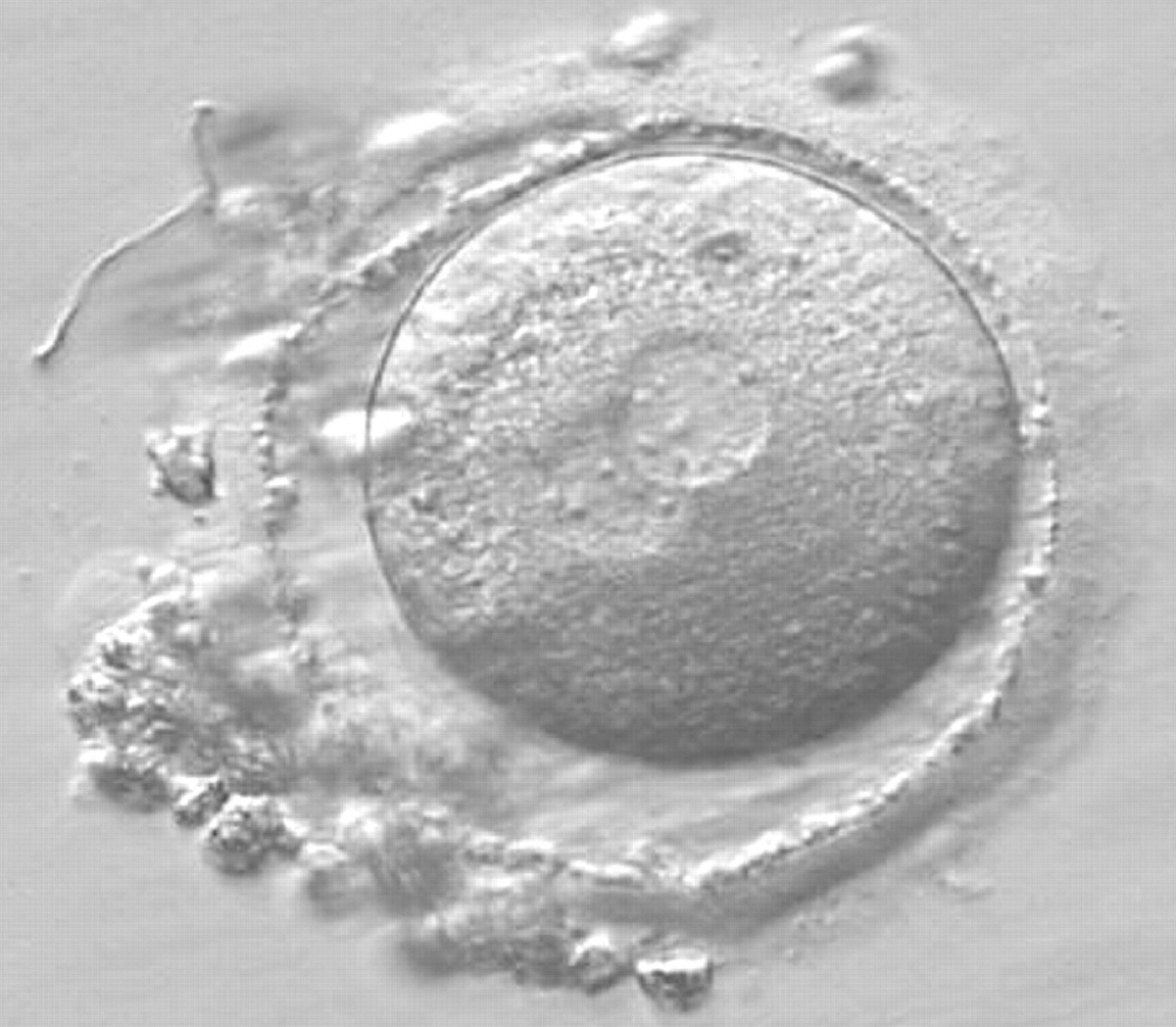
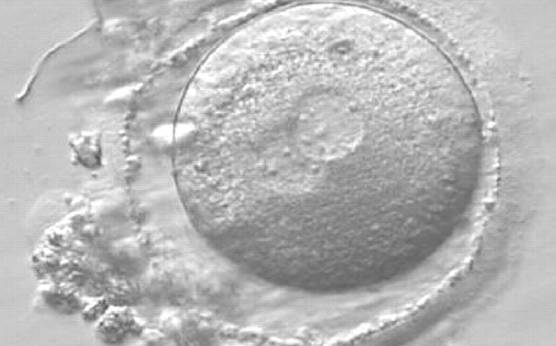
Figure 162
A zygote observed 16 h post-ICSI with small-sized NPBs scattered with respect to the PN junction in both PNs (400× magnification). It has a very thin ZP. It was transferred but failed to implant.
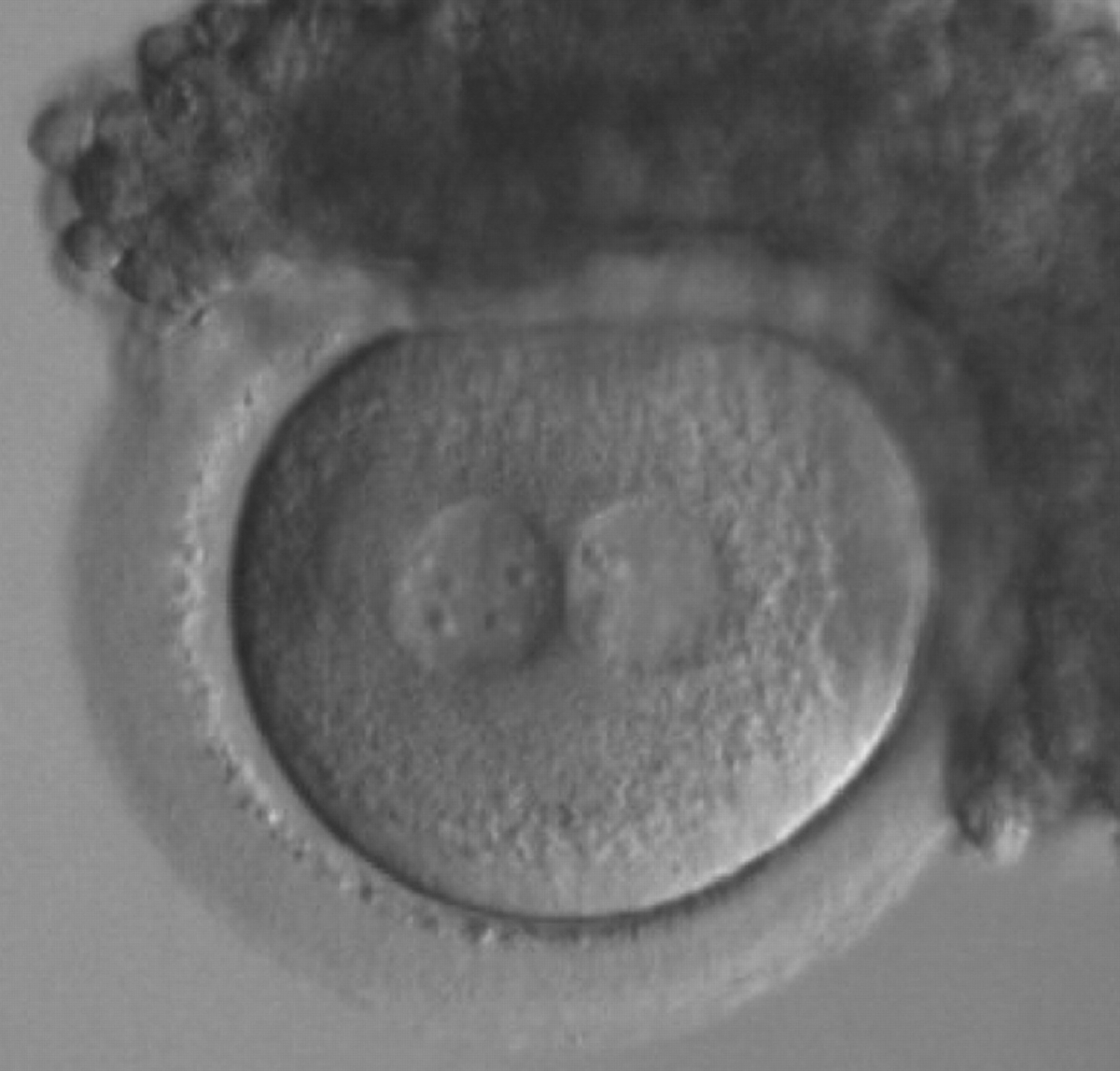
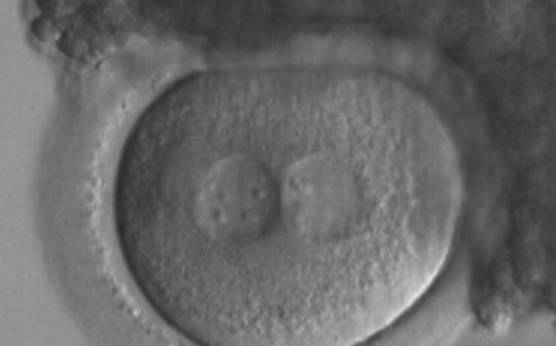
Figure 163
A zygote generated by ICSI with inequality in numbers and alignment of NPBs (200× magnification). Many granulosa cells are adherent to the ZP. It was transferred and implanted.
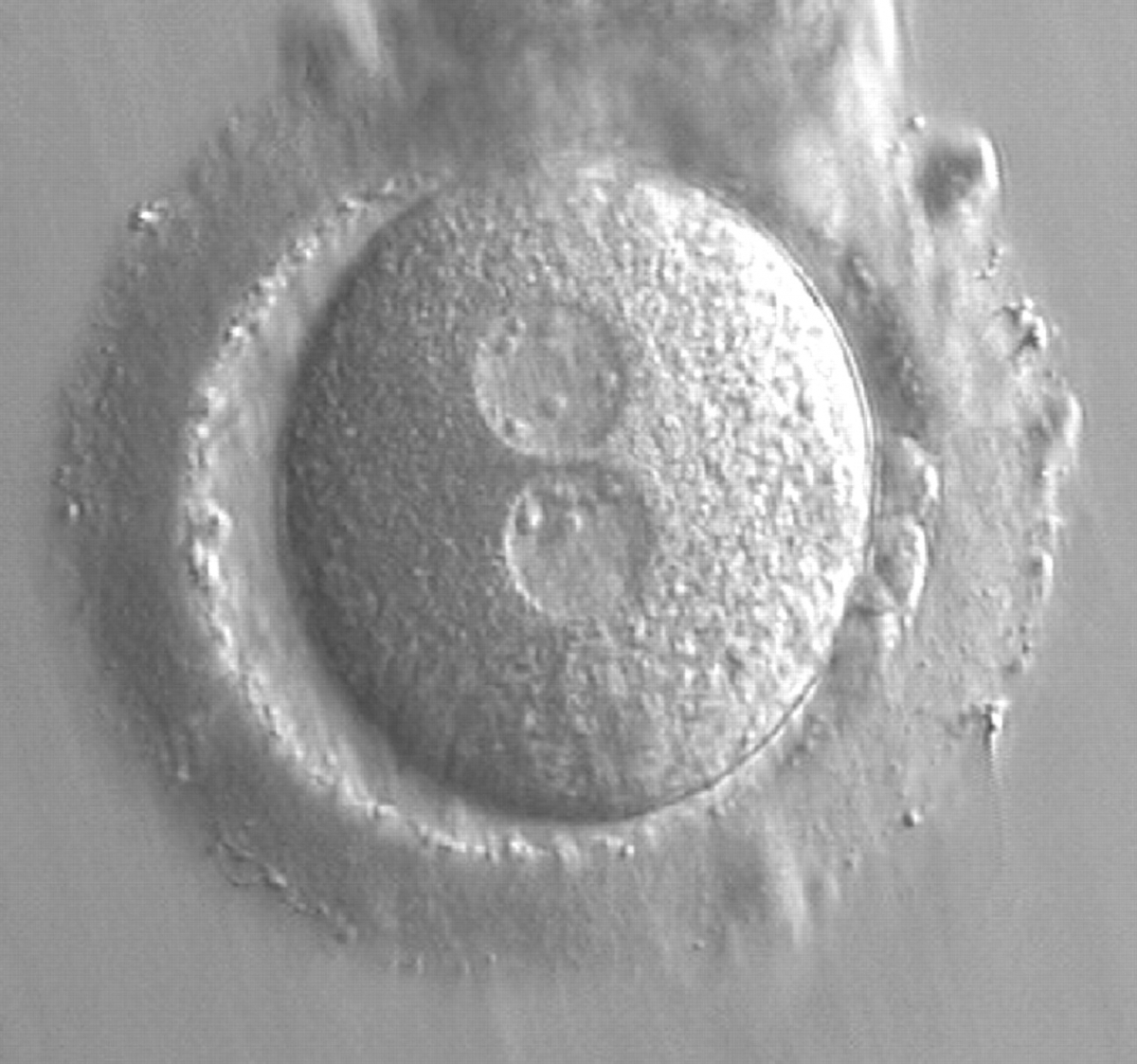
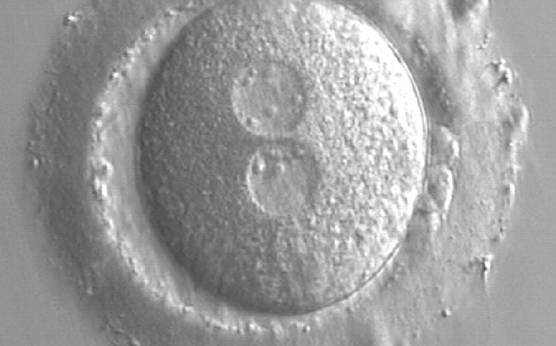
Figure 164
A zygote at 15 h post-IVF in which NPBs are scattered in one of the two PNs and aligned in the other (400× magnification). The ZP is thick and the PVS is enlarged. One of the two polar bodies is fragmented. It was transferred and implanted.
Pronuclear scoring based on Scott's (2003) Z scores, which combines the assessment of PN orientation and NPB pattern, was adopted by the consensus workshop (Alpha Scientists in Reproductive medicine and ESHRE Special Interest Group of Embryology, 2011) but modified into three categories:
- symmetrical PNs (corresponding to Scott's Z1 and Z2)
- non-symmetrical PNs (other arrangements, peripheral PNs)
- abnormal PNs (PNs with none or one NPB, which are called respectively ‘ghost’ PNs and ‘bulls-eye’ PNs. Both have been associated with abnormal outcome in animal models.
D.2 Numbers similar/numbers different between PNs
Human cells generally have two to seven nucleoli per human nucleus with equal numbers in the two daughter cells after a mitotic division. Nucleoli appear and disappear depending on the cell cycle phase: they are more numerous at the G1 phase, then start to fuse, and at the S1 phase there are only one to two large nucleoli per nucleus. When asynchrony occurs, this appears to be the result of aberrant chromosomal function.
Transferring this model to the zygote, the ideal oocytes are those presenting with symmetry for number and size of NPBs that are aligned on the furrow between the 2PNs (Figs 165–167). Equality in number with non-alignment in both PNs is also indicative of synchronised development (Fig. 168). In contrast, any form of disparity in NPBs' size, number or pattern of alignment between the PNs, is associated with a poor outcome (Figs 169–172).
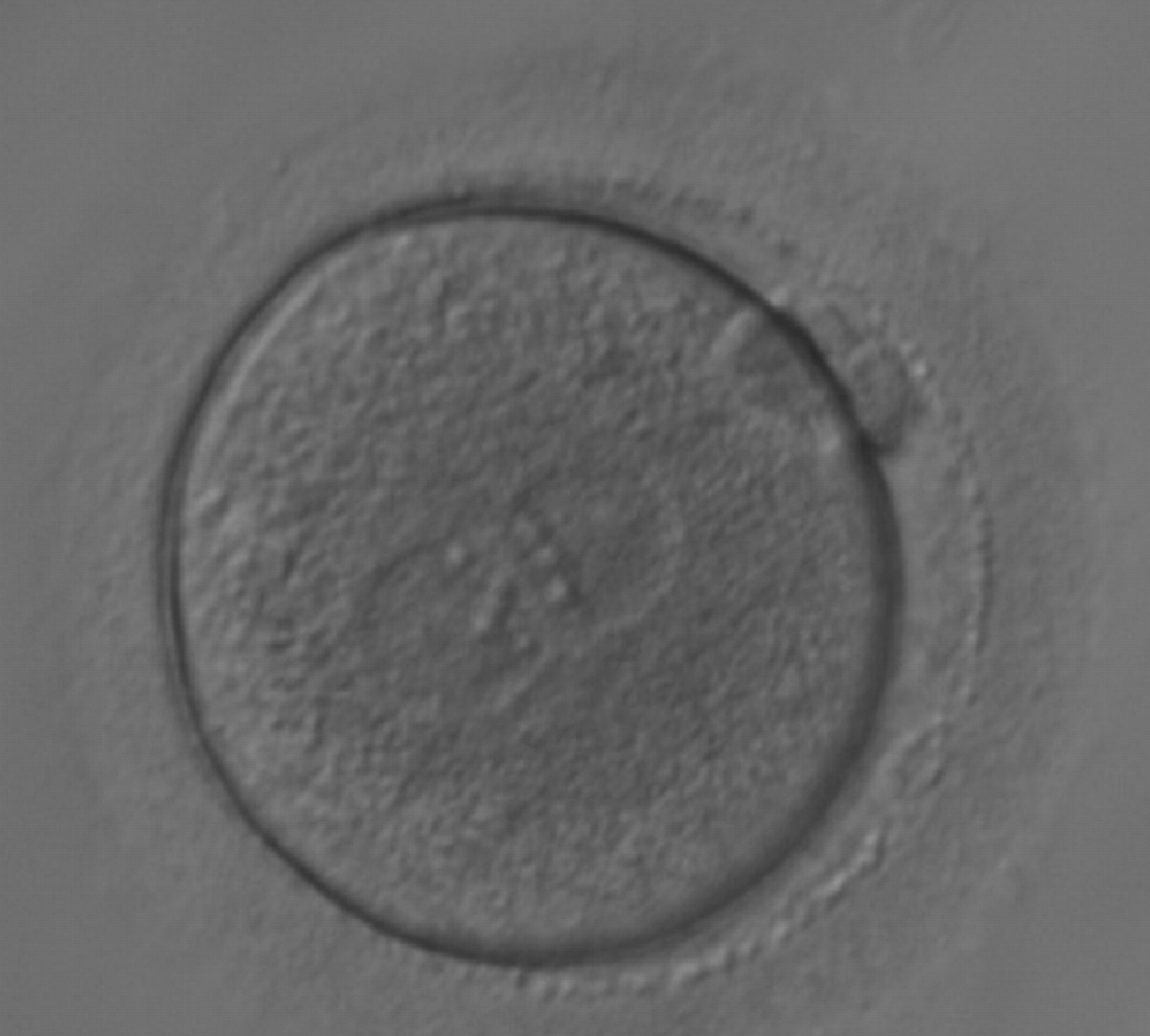
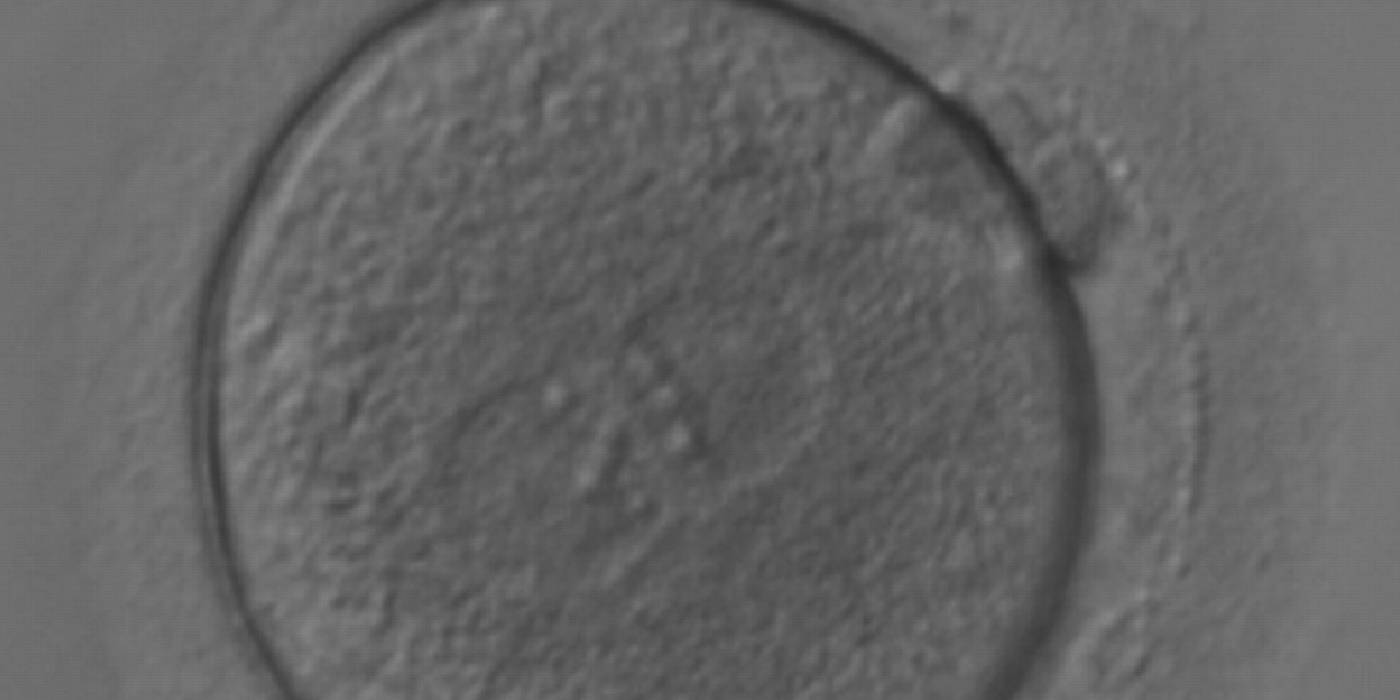
Figure 165
A zygote generated by ICSI with equal numbers of large-sized NPBs aligned at the PN junction (200× magnification). It was transferred but the outcome is unknown.

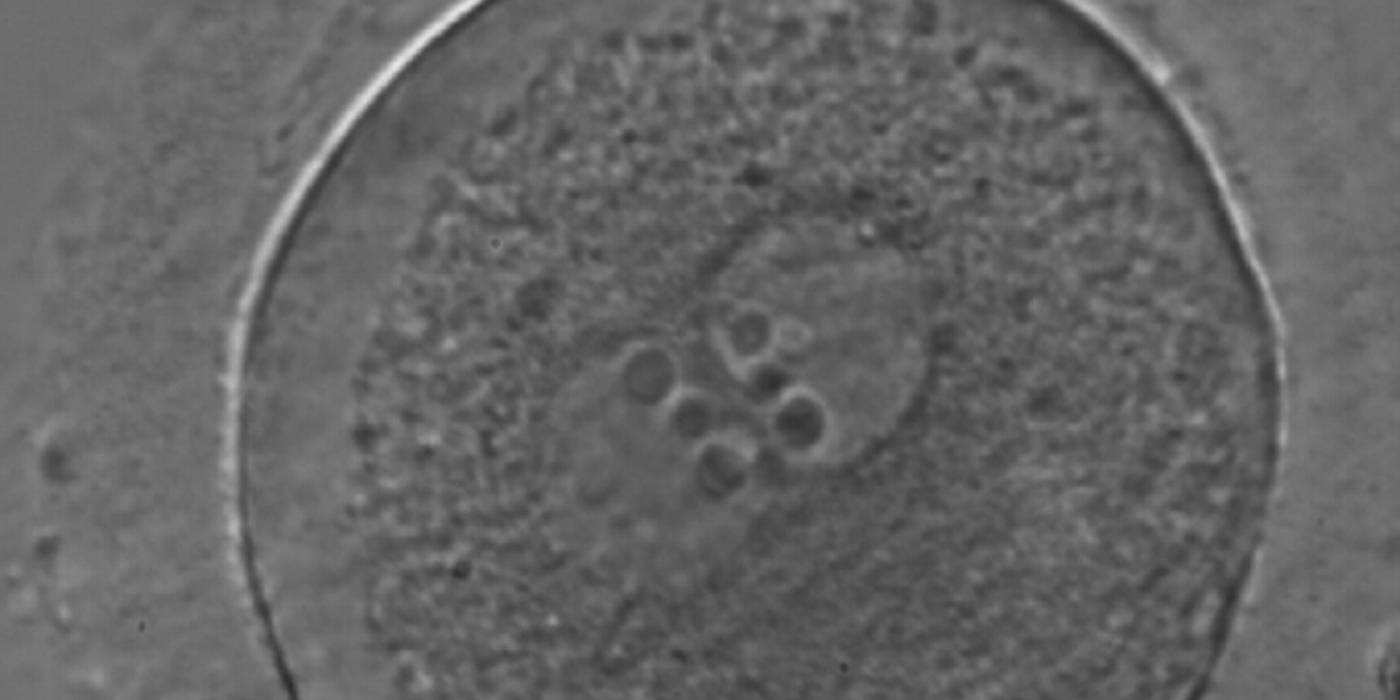
Figure 166
A zygote generated by ICSI with equal numbers of large-sized NPBs aligned at the PN junction (400× magnification). There is a halo in the cortical area; polar bodies are fragmented and the ZP appears brush-like. It was discarded due to subsequent abnormal development.
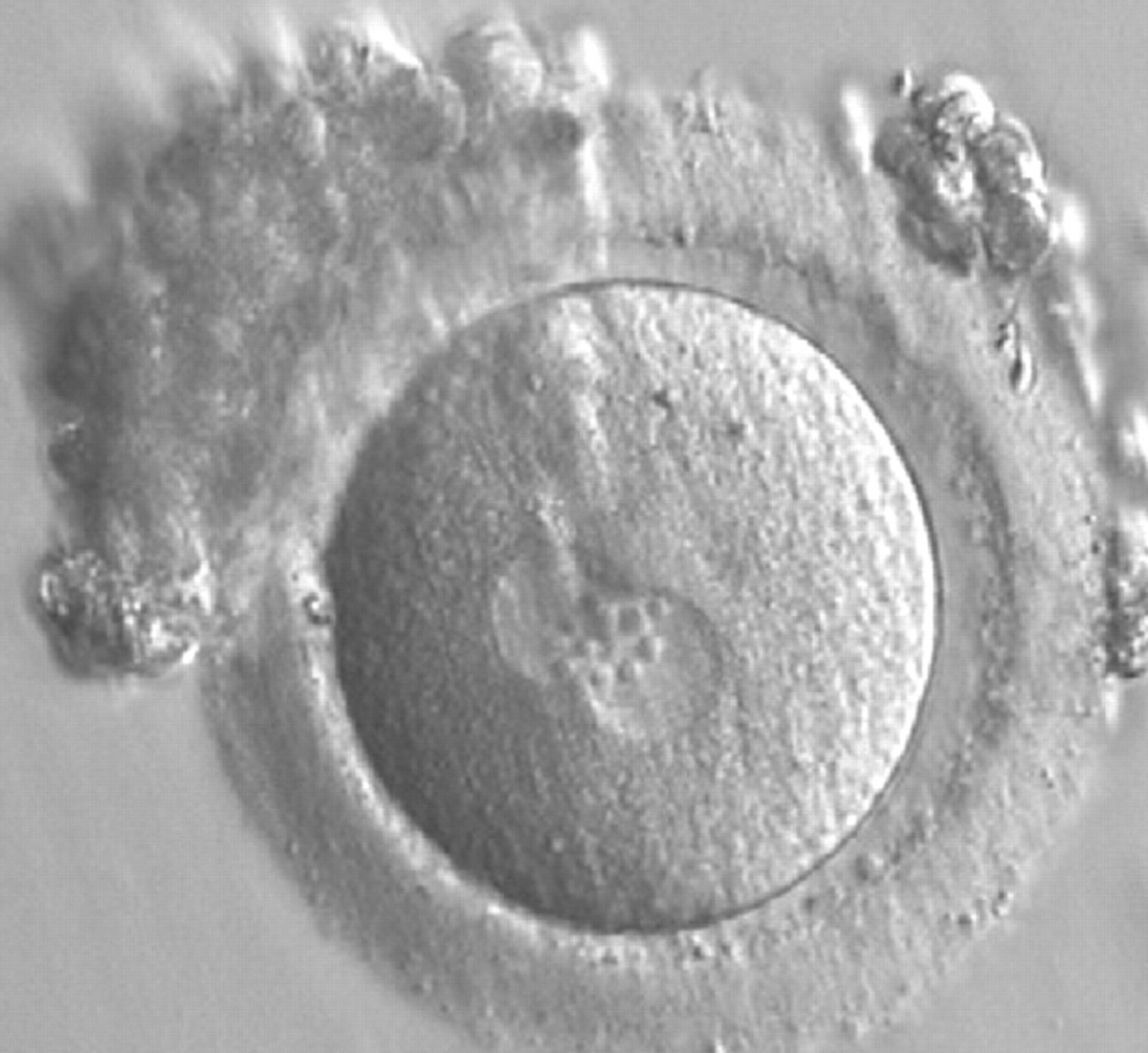
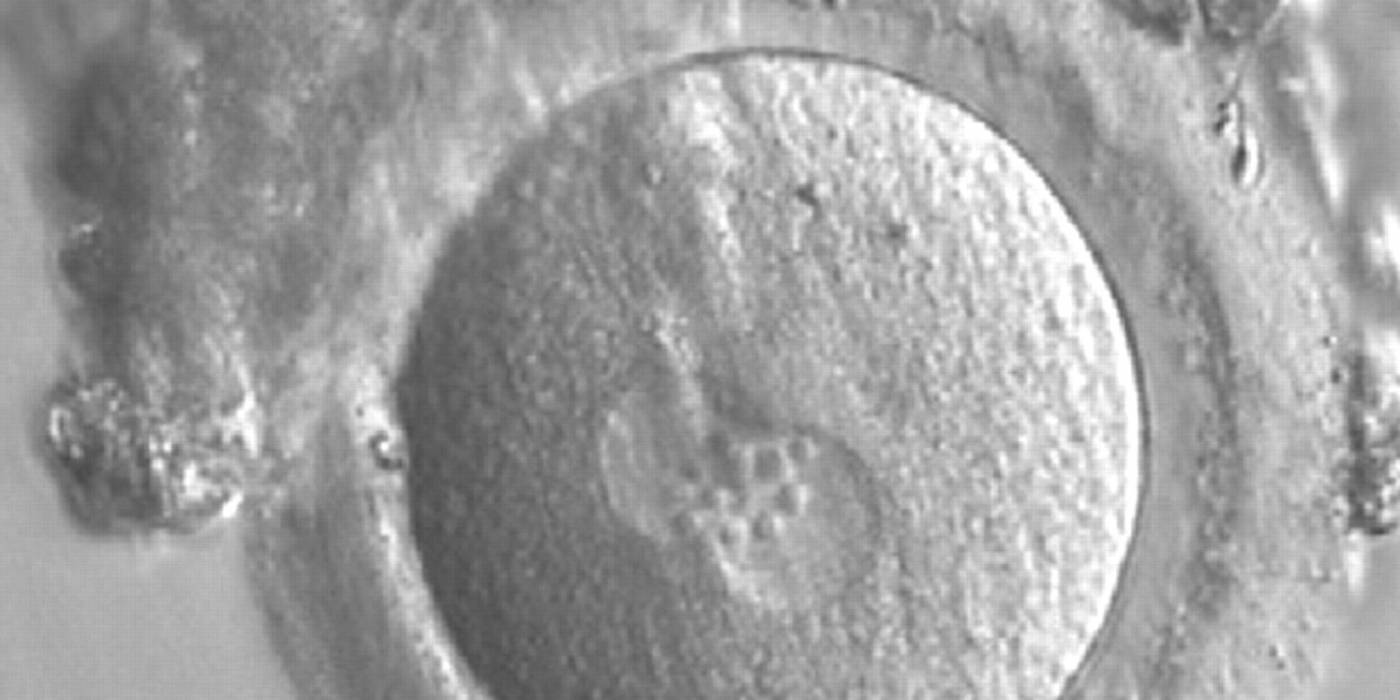
Figure 167
A zygote generated by IVF observed 17 h post-insemination (400× magnification). PNs have similar numbers of large-sized NPBs aligned at the PN junction. The PVS is slightly enlarged and the ZP is normal in size and surrounded by some granulosa cells. It was transferred but failed to implant.
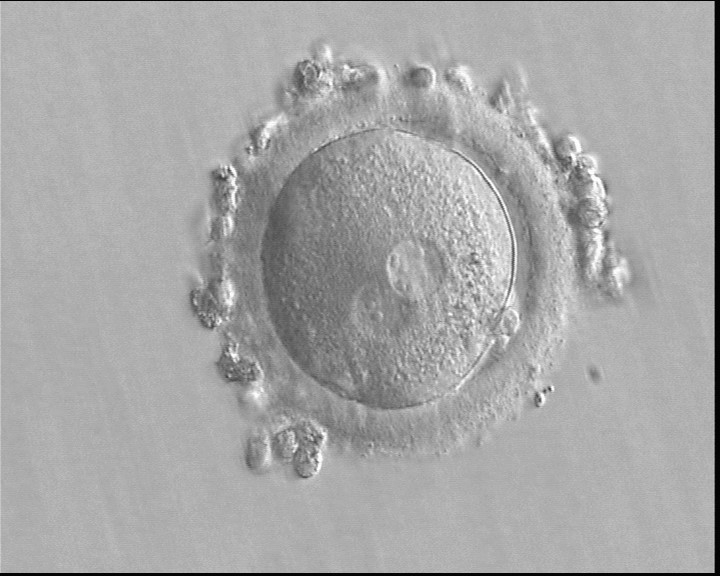
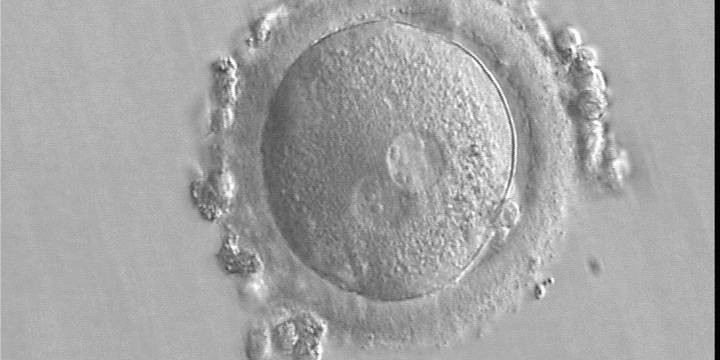
Figure 168
A zygote generated by IVF using frozen/thawed ejaculated sperm and observed 15 h post-insemination (400× magnification). Two PNs of approximately the same size are clearly visible in the cytoplasm. Peripheral granular cytoplasm can be seen. NPBs are scattered in both PNs. Both polar bodies are located at the 4 o'clock position. It was transferred and implanted.
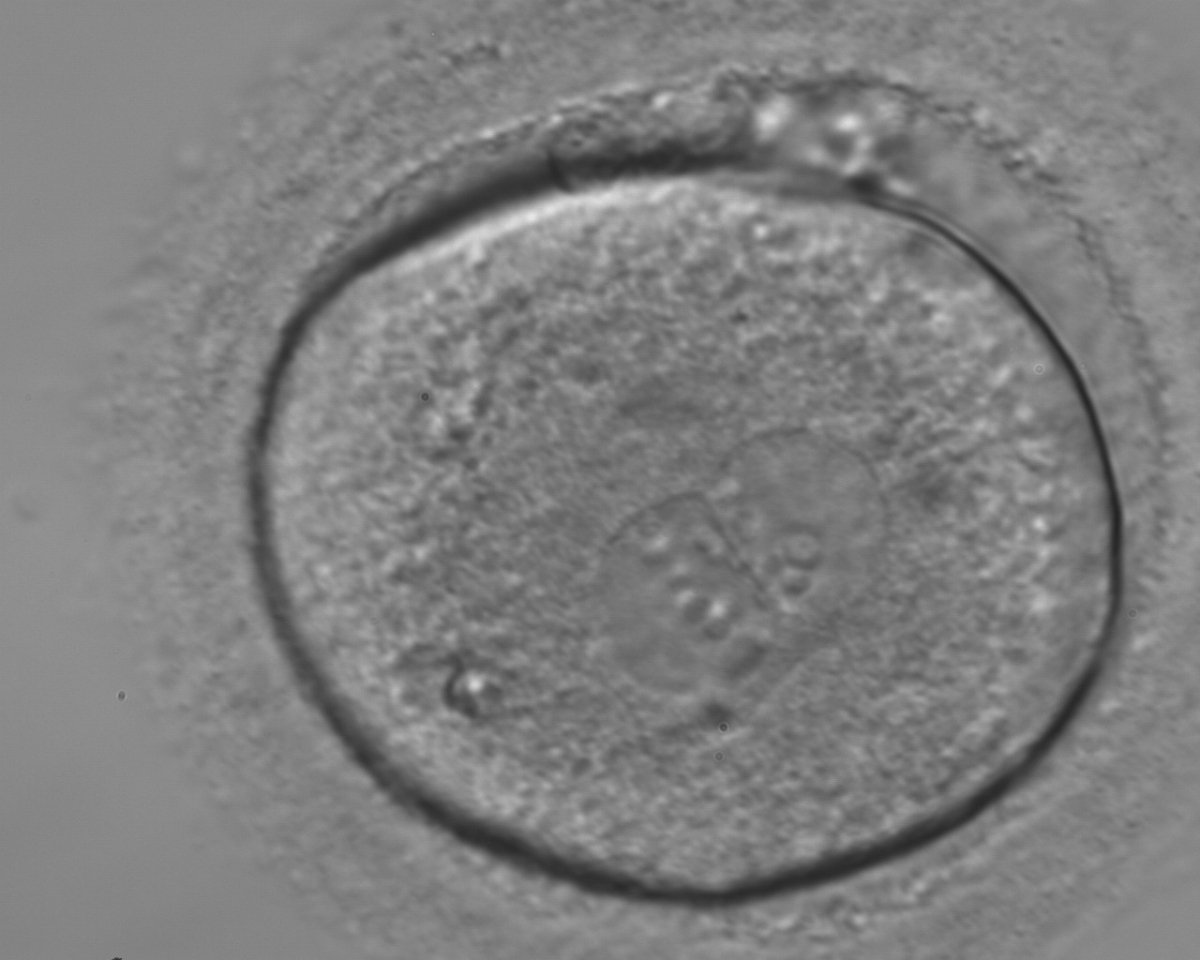
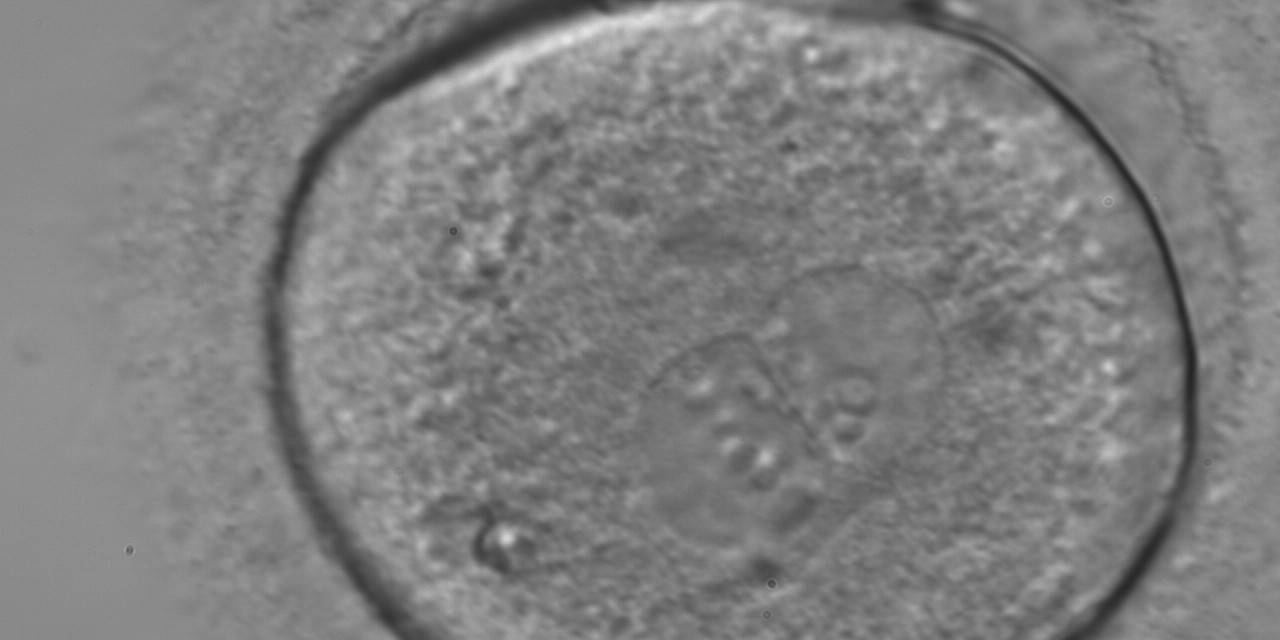
Figure 169
A zygote generated by ICSI showing different numbers of NPBs (400× magnification). The cytoplasm appears granular. It was transferred but failed to implant.
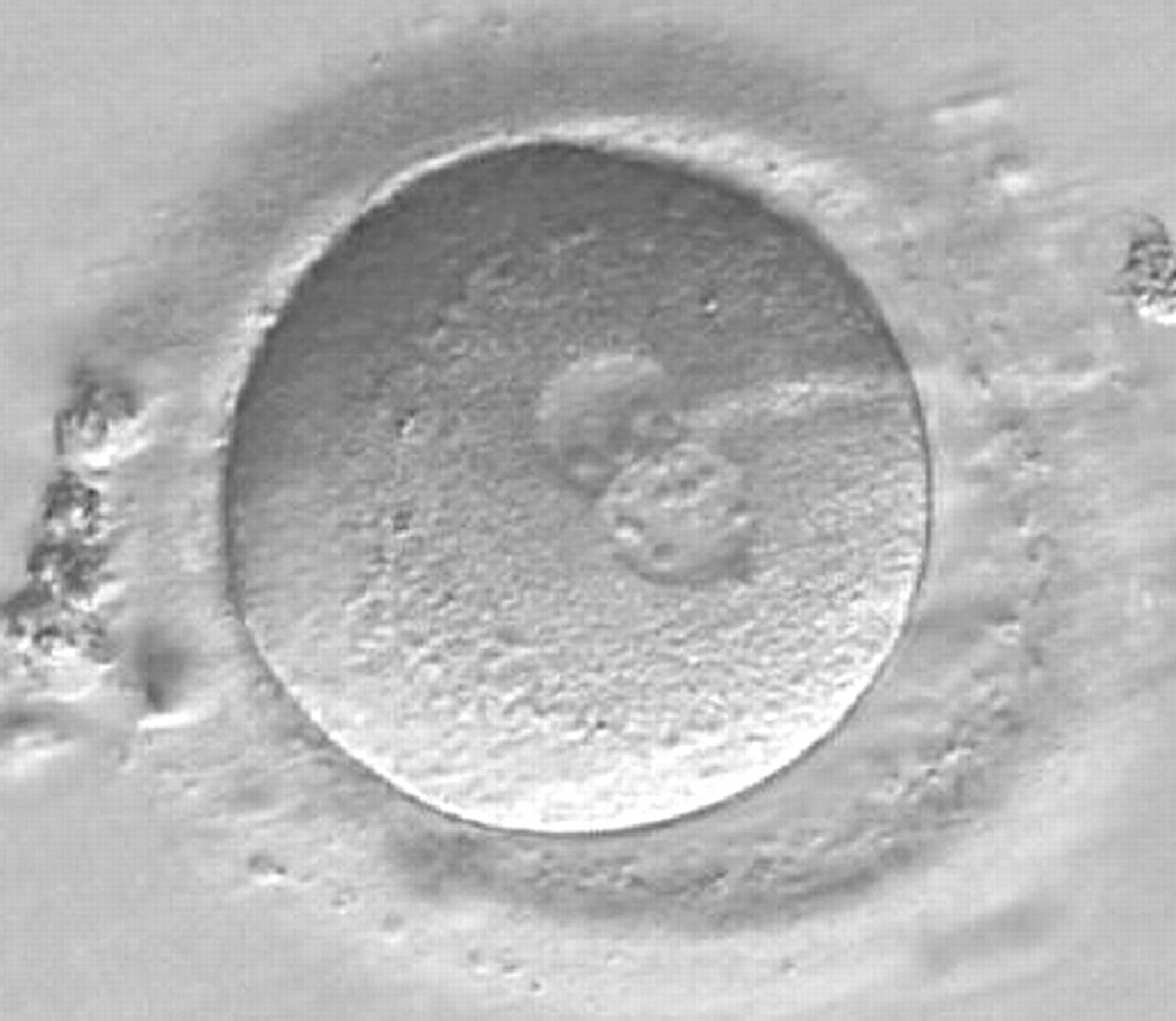
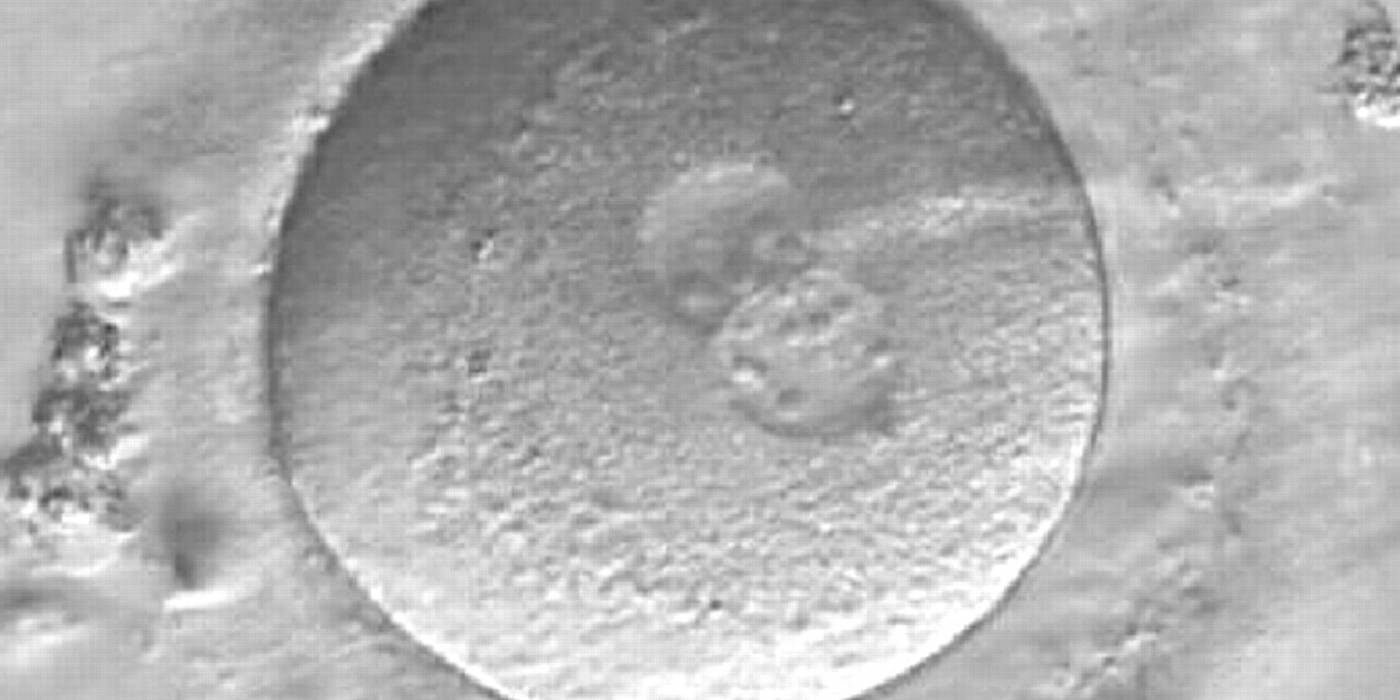
Figure 170
A zygote observed 16.5 h post-ICSI, showing unequal number and size of NPBs: medium-sized and scattered in one PN, larger-sized and aligned in the other (400× magnification). Polar bodies had been biopsied, and the slit opened mechanically in the ZP is evident at the 3 o'clock position. It was cryopreserved.
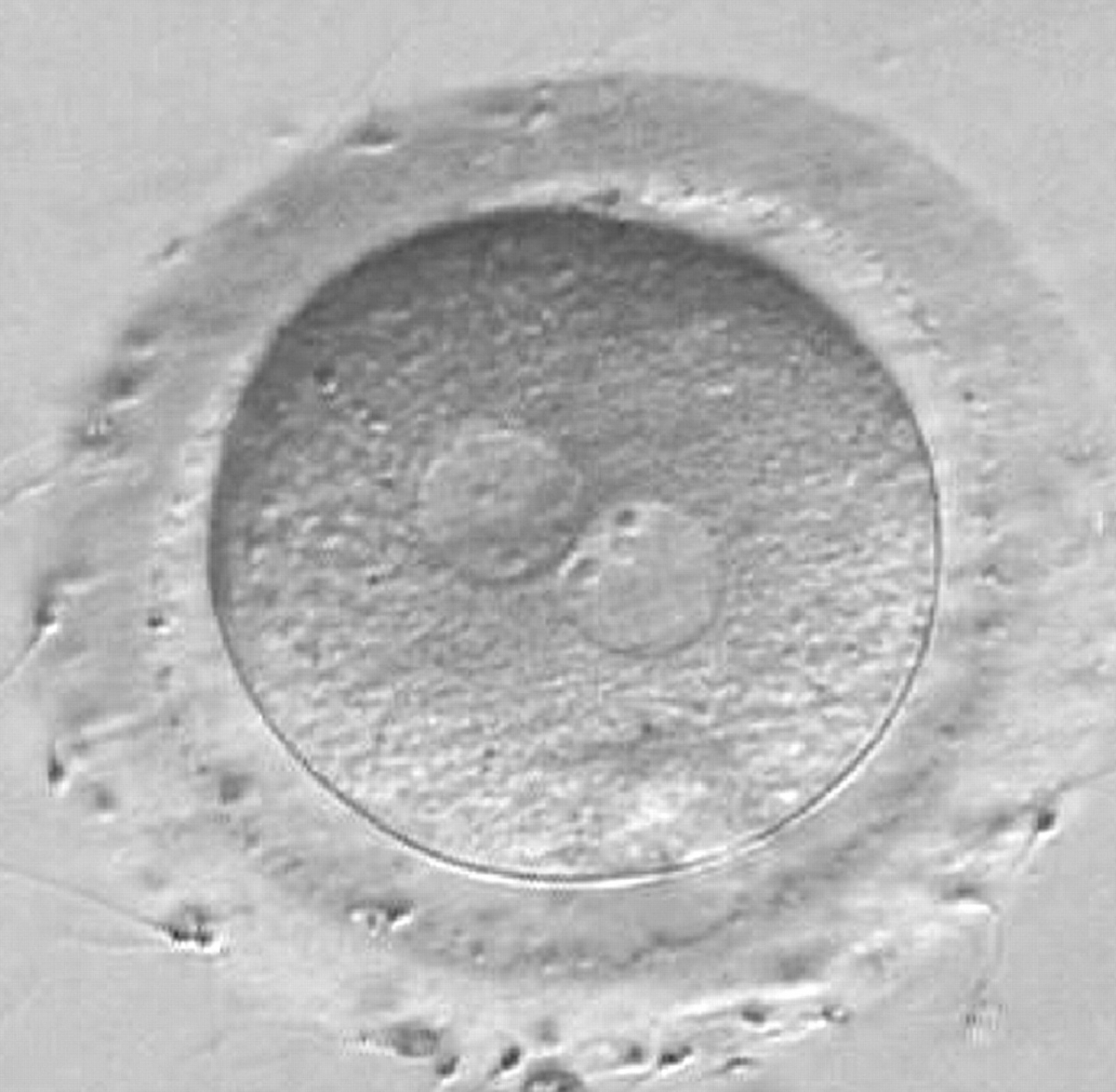
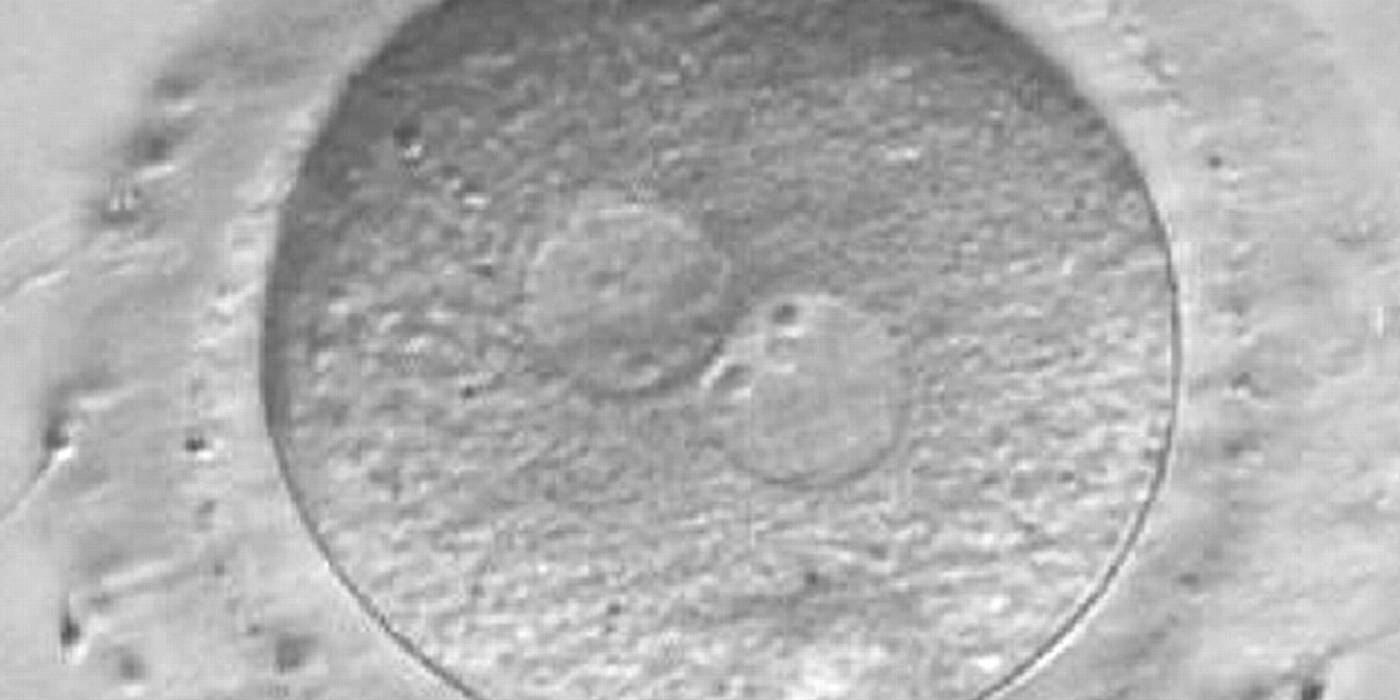
Figure 171
A zygote observed at 16 h post-IVF displaying different number and size of NPBs: small and scattered in one PN, large and aligned in the other (400× magnification). It was discarded because of developmental arrest.
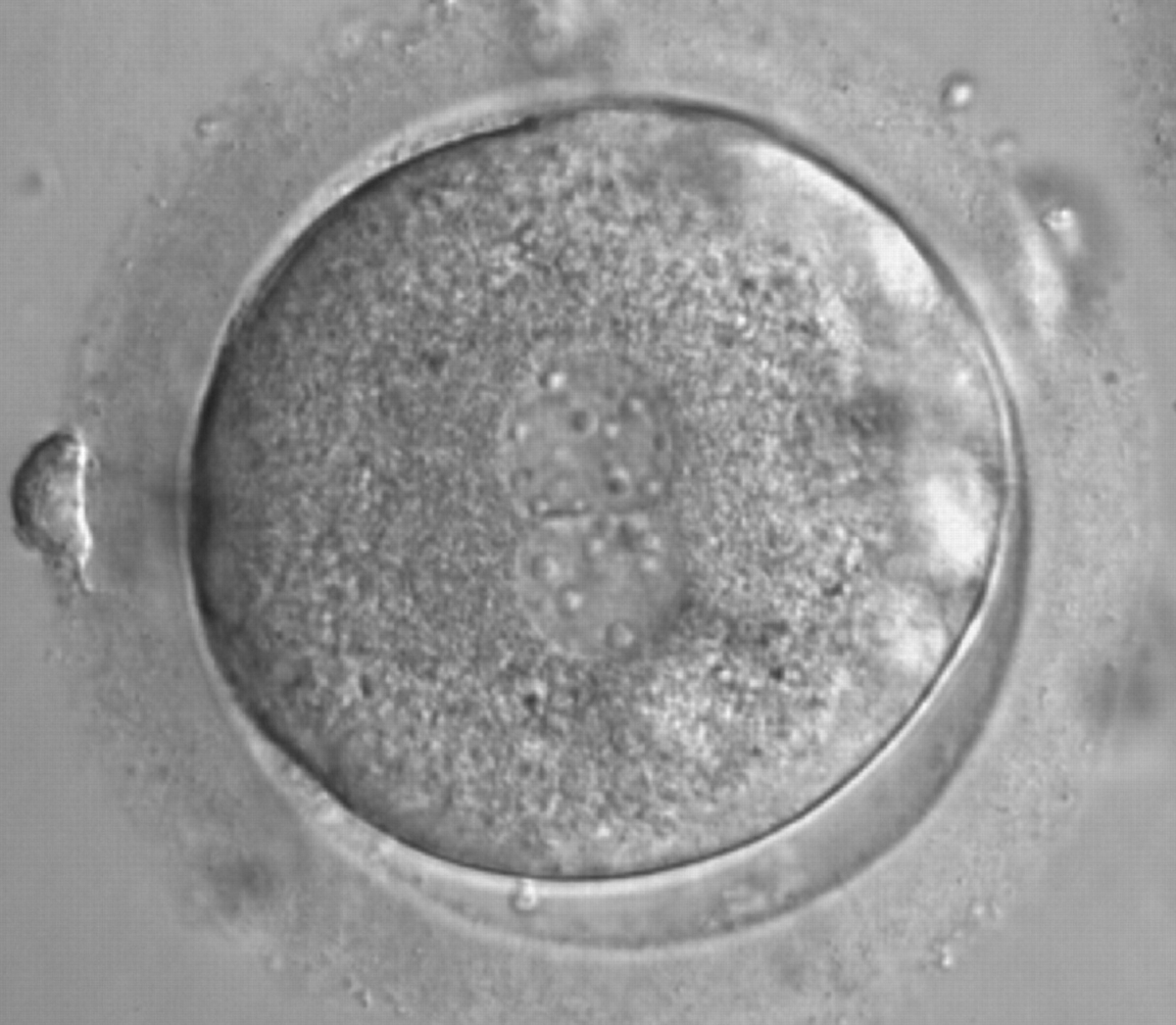
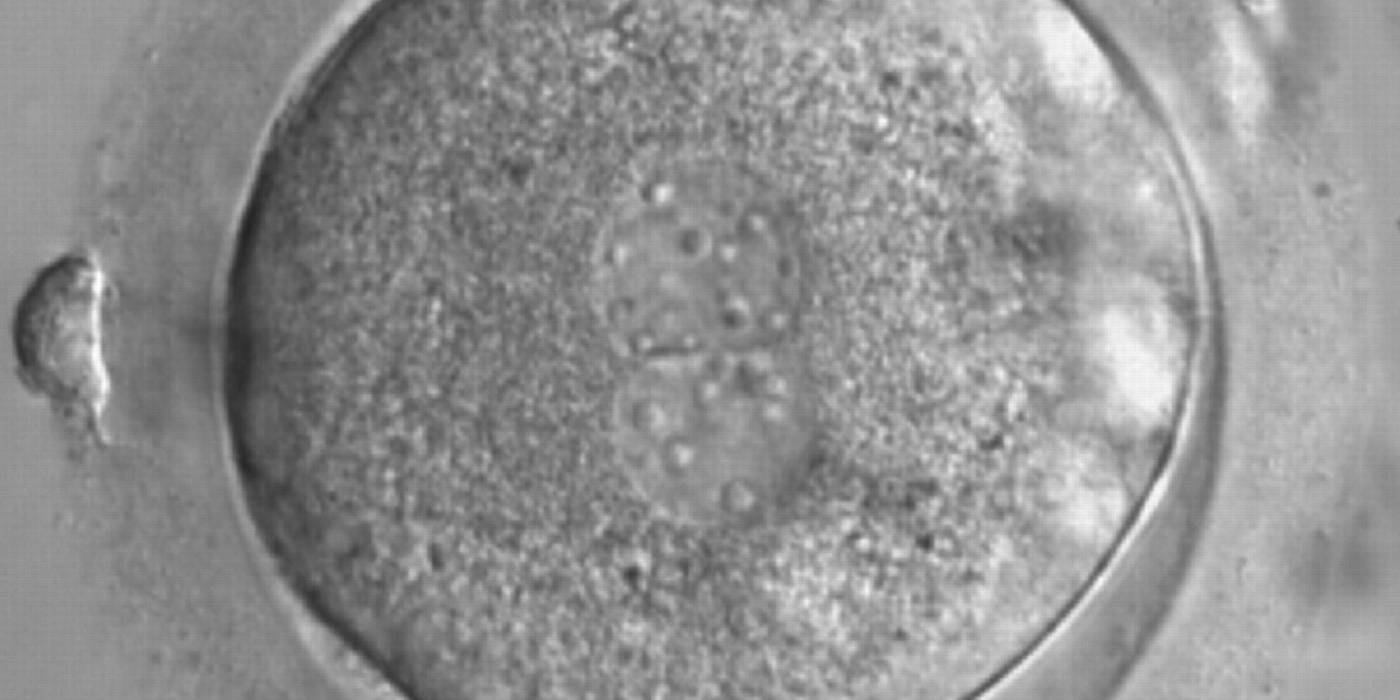
Figure 172
A zygote generated by ICSI with a different number and size of scattered NPBs (600× magnification). It was discarded because of subsequent abnormal development.
D.3 Similar size of NPBs/different size of NPBs
The dynamics of nucleoli formation associated with NPBs merging implies a time-related change in NPBs' size (Scott, 2003). A progressive increase in NPBs' size and a concomitant reduction in number occur at the time of nucleoli alignment into the furrow between PNs (Figs 173–176).
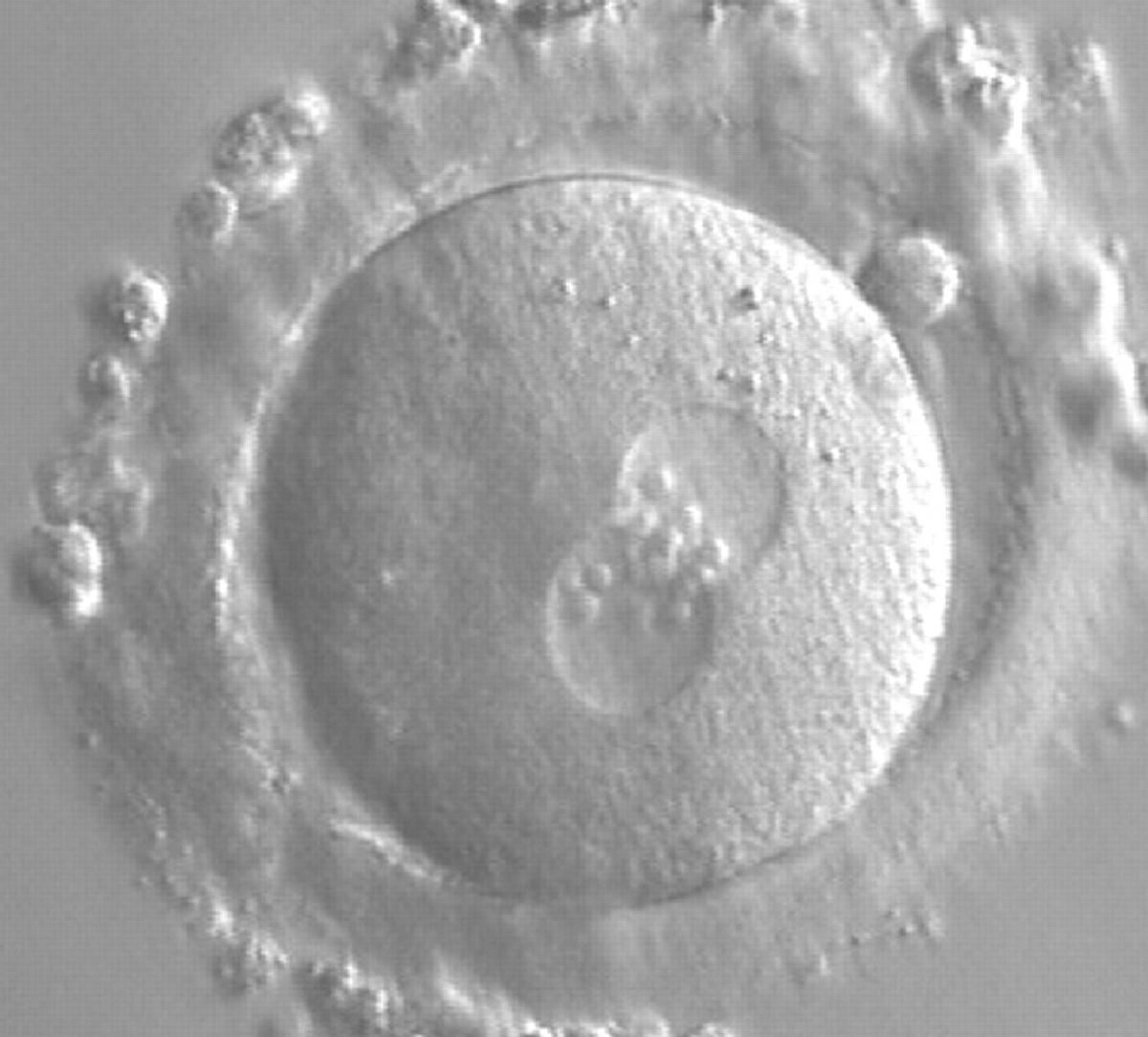
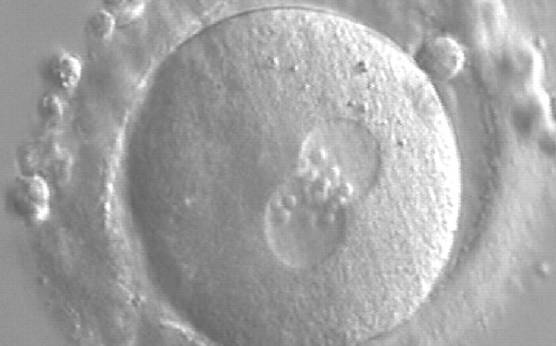
Figure 173
A zygote generated by ICSI showing equal number and size of NPBs, which are aligned at the PN junction (200× magnification). PNs are tangential to the plane of the polar bodies. It was transferred and resulted in a singleton pregnancy and delivery.
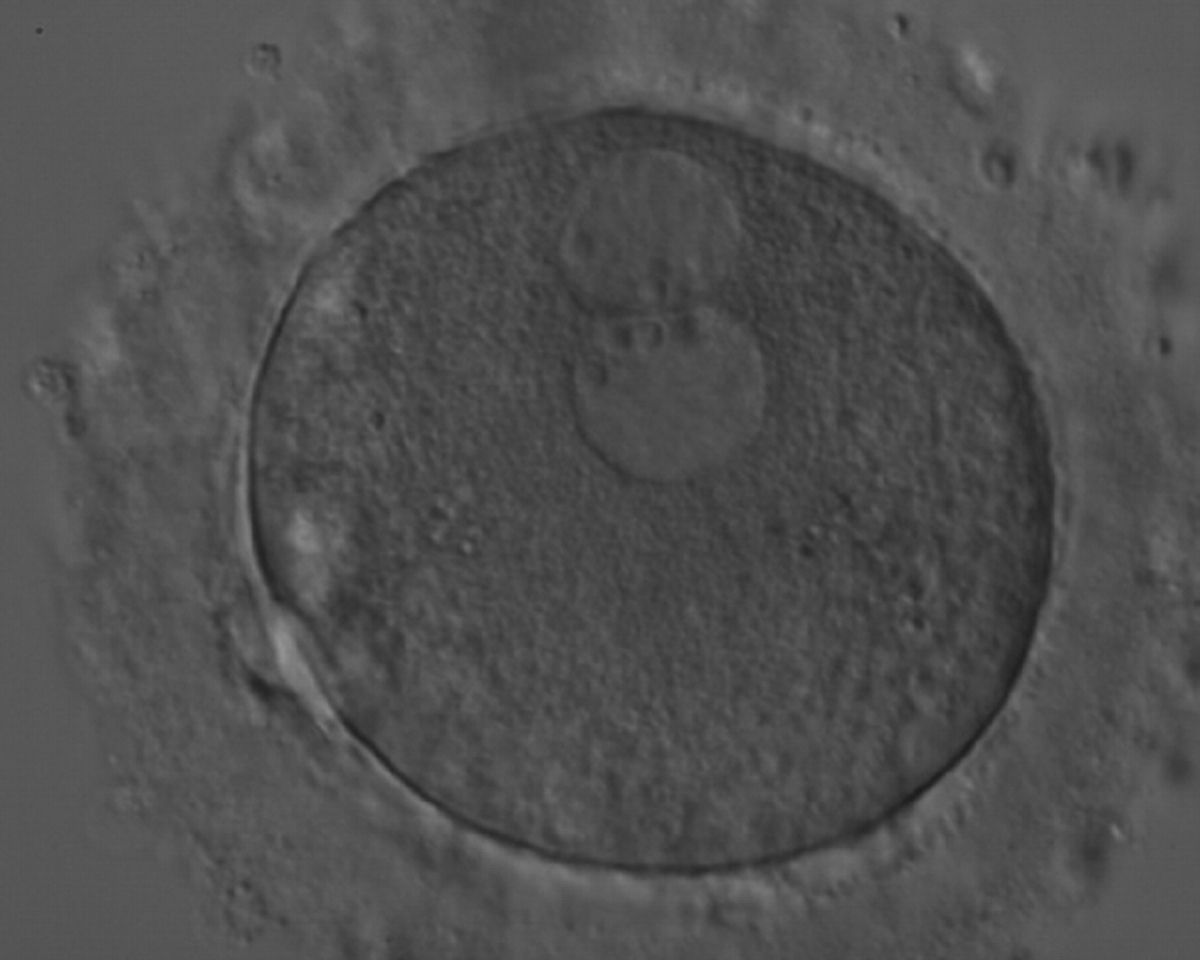
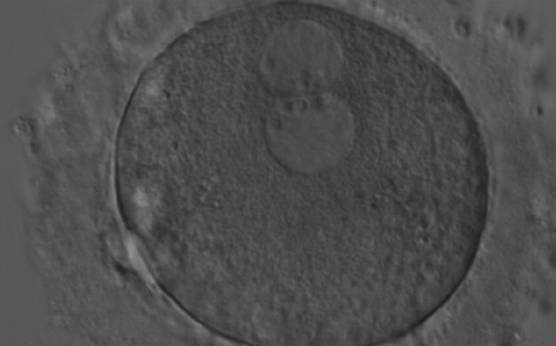
Figure 174
A zygote generated by ICSI with peripherally located PNs showing NPBs of similar size and number (200× magnification). Further development resulted in uneven cleavage and arrest on Day 3.
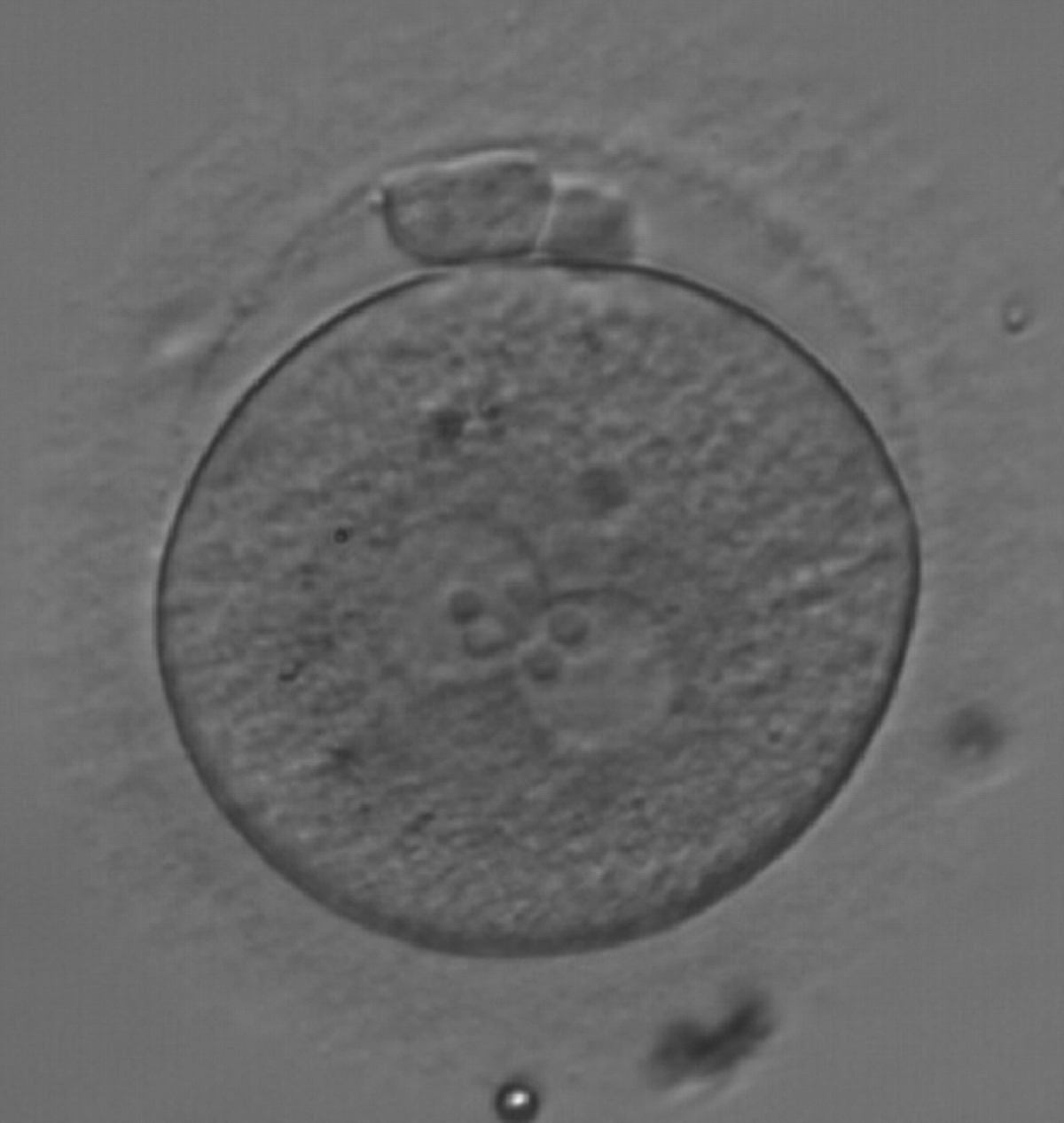
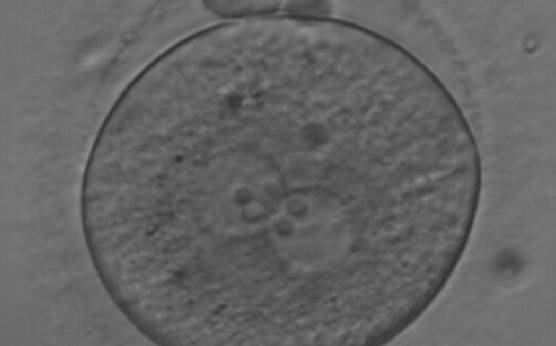
Figure 175
A zygote observed at 18 h after ICSI, with NPBs of equal size in both PNs and aligned at the PN junction (200× magnification). Polar bodies are intact and slightly larger than normal. The cytoplasm is granular with some inclusions. It was discarded due to subsequent abnormal development.
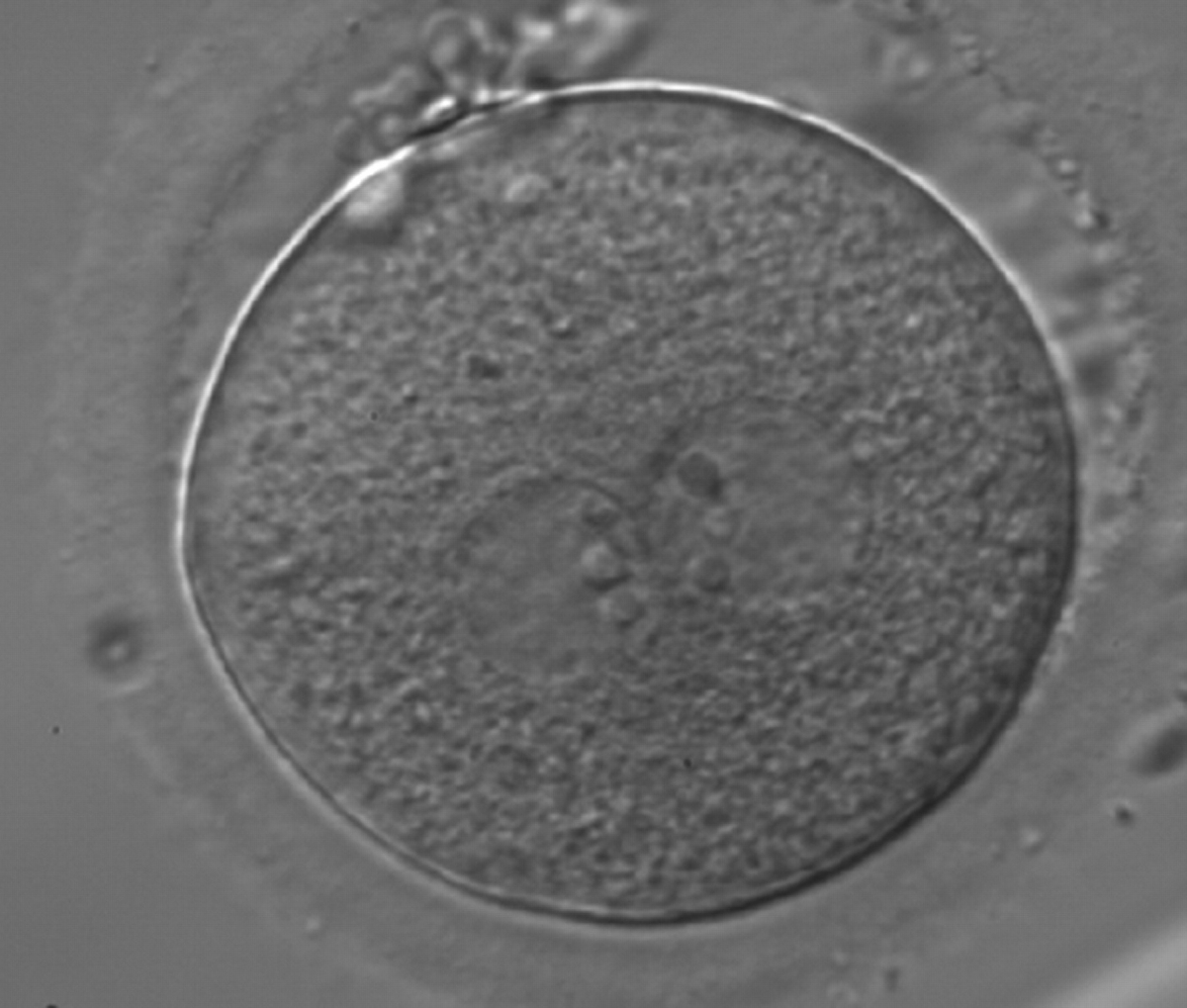
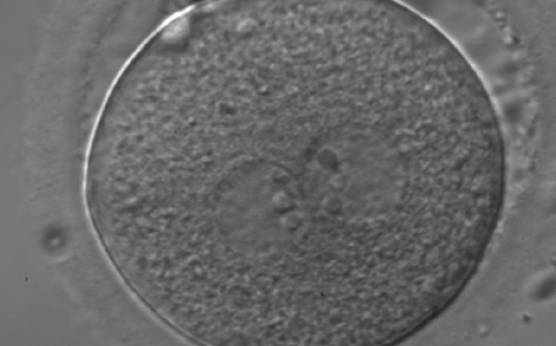
Figure 176
A zygote generated by ICSI showing equal number and size of NPBs, which are perfectly aligned at the PN junction (200× magnification). Polar bodies are highly fragmented. It was transferred but clinical outcome is unknown.
The presence of small scattered and unequal-sized NPBs (Figs 177–179) could be indicative of functional defects in nucleoli with consequent decreased or ineffective synthesis of rRNA. The extent of malfunctioning possibly depends on the grade of asynchrony and on the number of affected nucleoli as demonstrated by the clinical outcome that may result in implantation (Fig. 180).
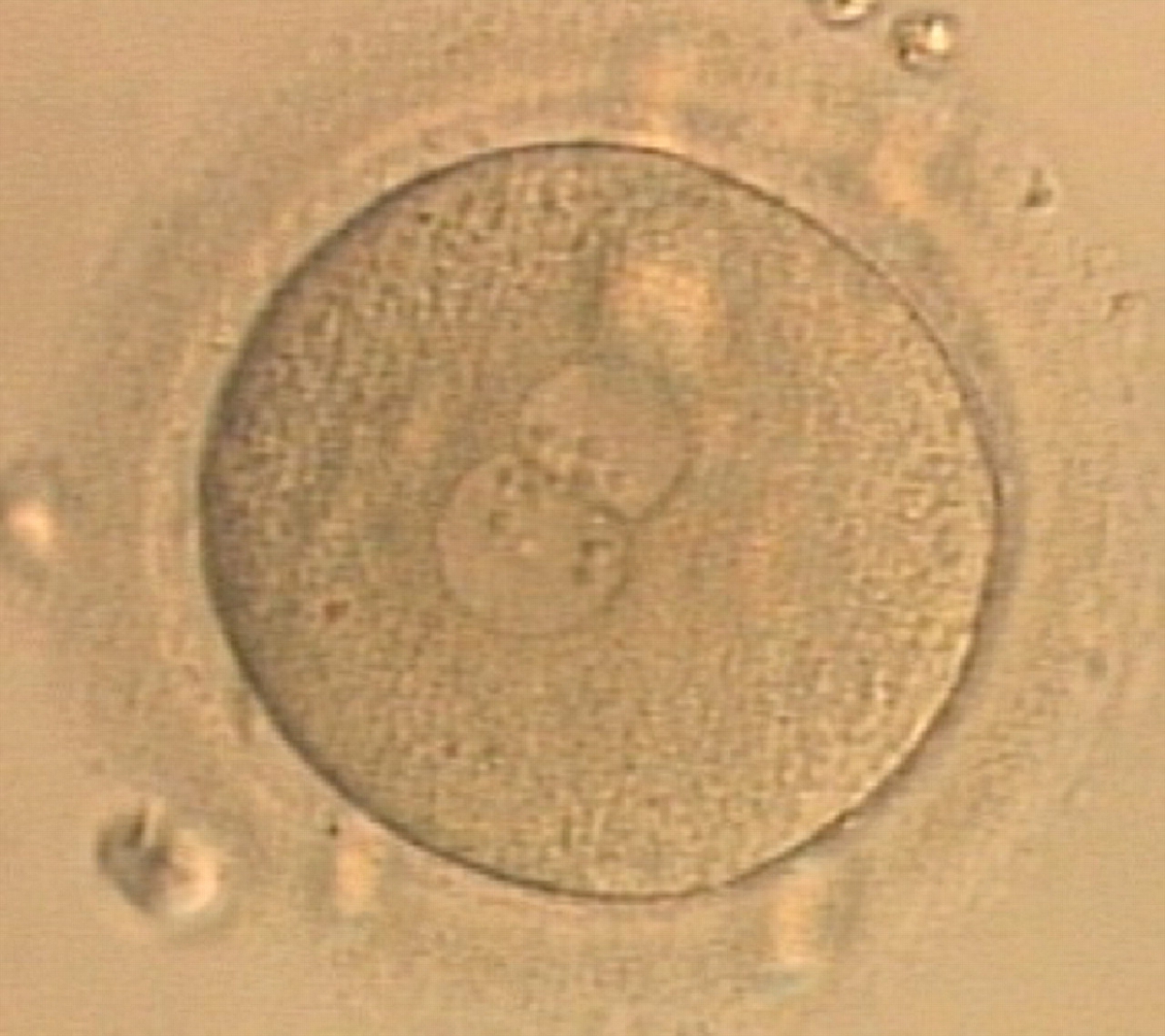
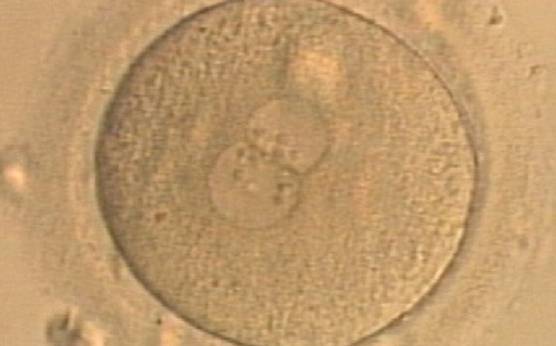
Figure 177
A zygote generated by ICSI displaying unequal number and size of NPBs between the PNs (400× magnification). It was transferred but clinical outcome is unknown.
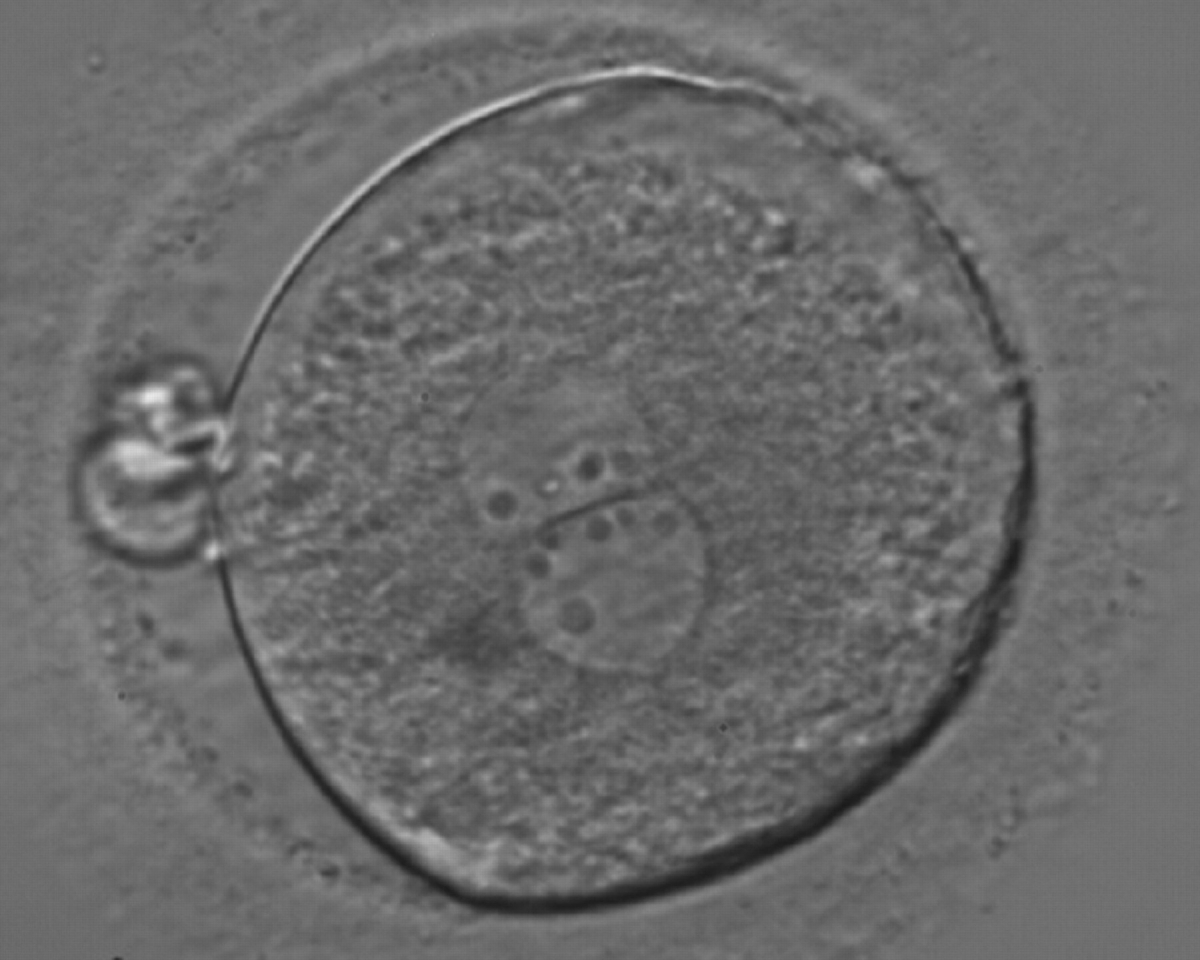
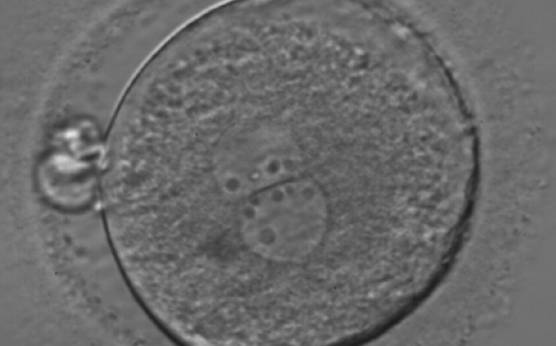
Figure 178
A zygote generated by ICSI showing an unequal number of NPBs (400× magnification). The NPBs differ in size within each PN. It was transferred but clinical outcome is unknown.
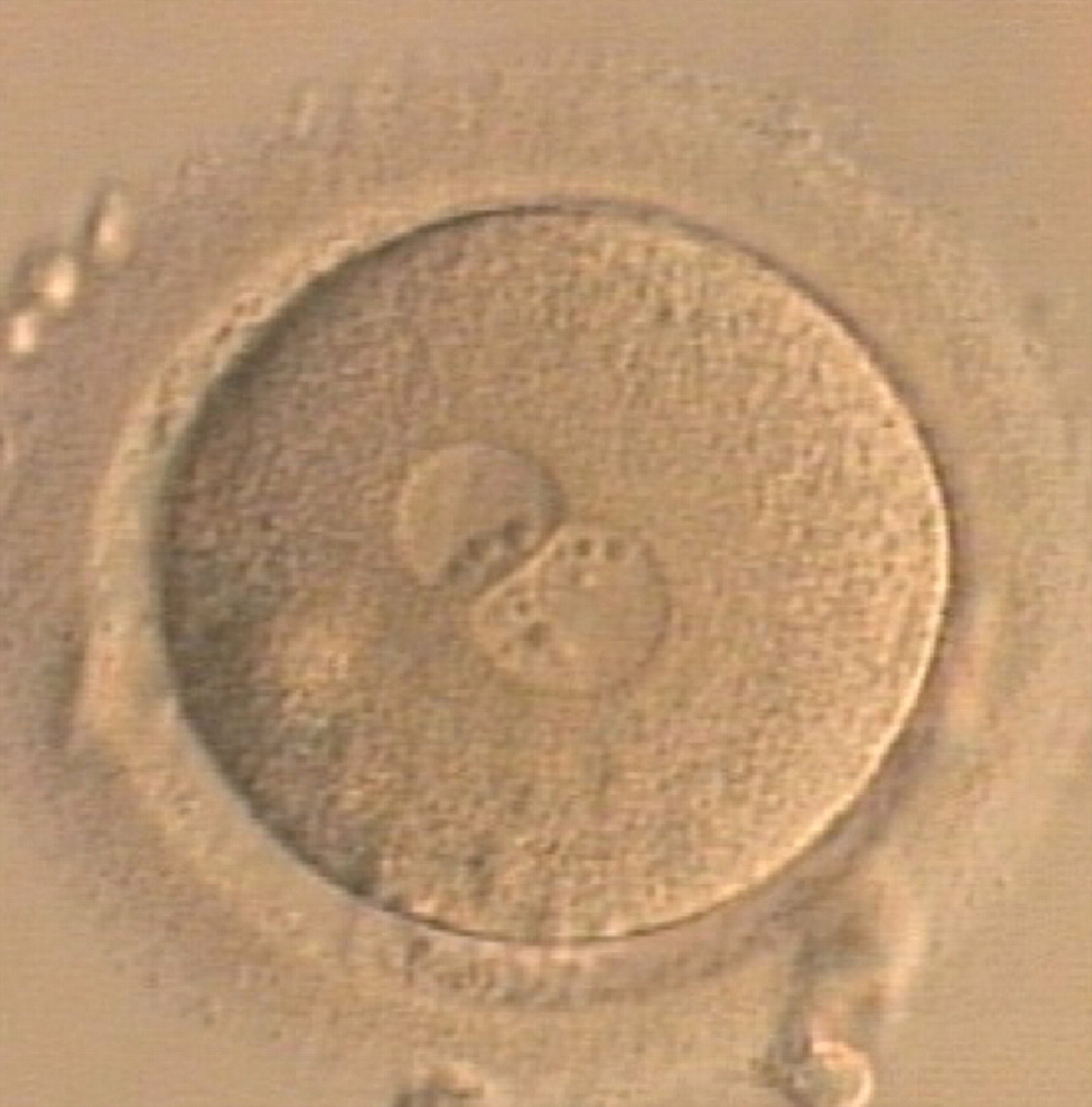
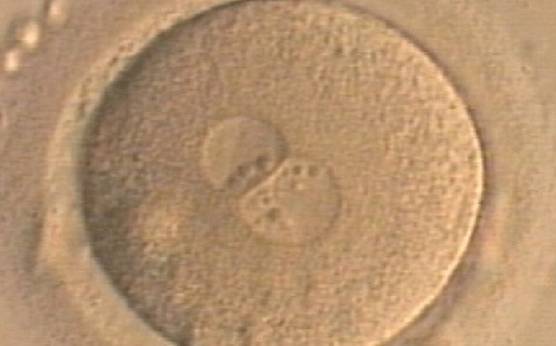
Figure 179
A zygote generated by ICSI displaying unequal number and size of NPBs between the PNs (400× magnification). NPBs are mainly aligned at the PN junction. It was transferred but clinical outcome is unknown.
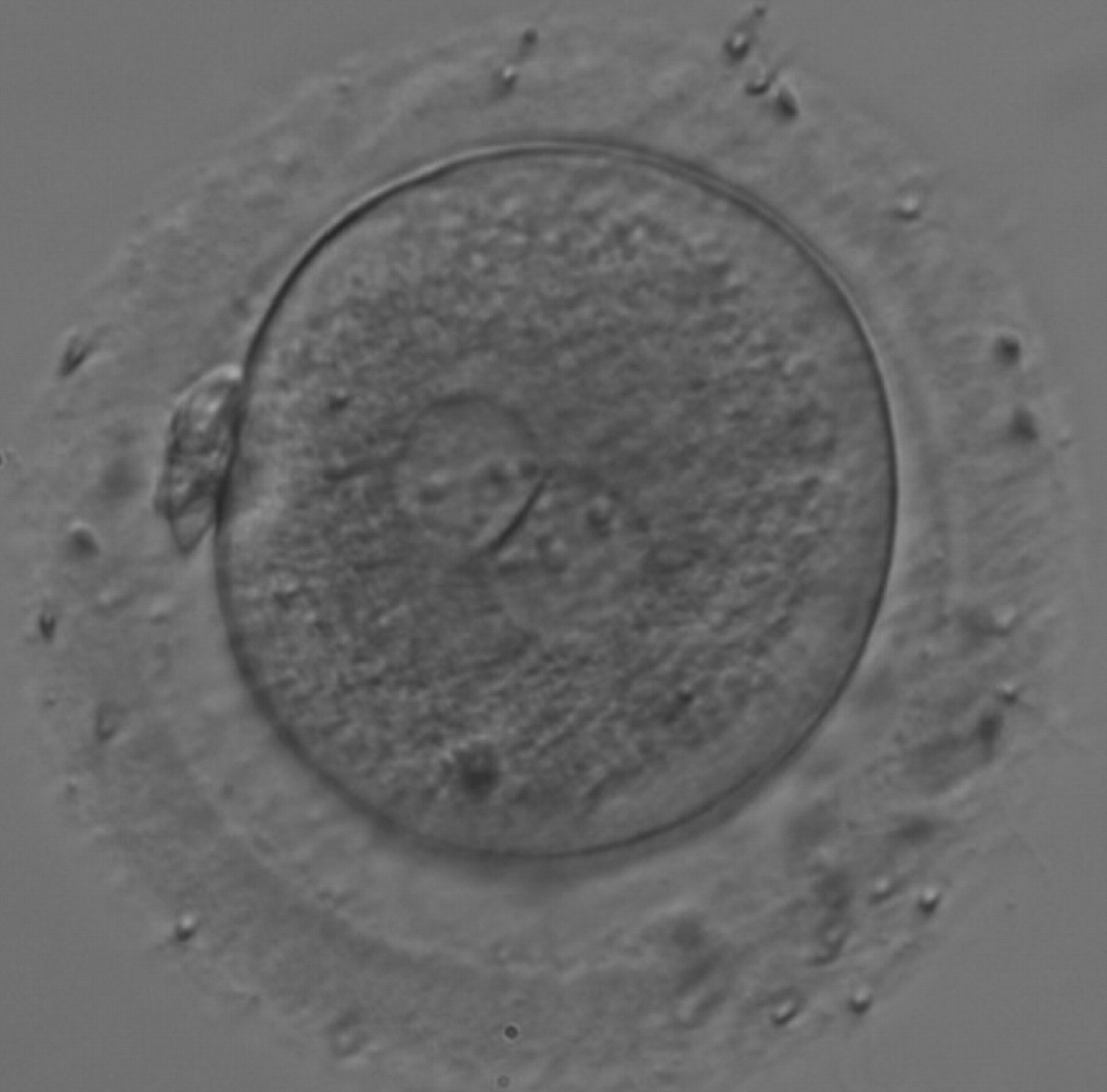
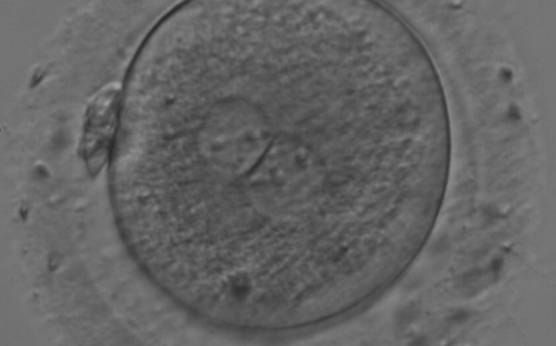
Figure 180
A zygote generated by IVF with large NPBs scattered with respect to the PN junction (200× magnification). NPBs display differences in sizes within each PN and are slightly larger in the PN on the right side. It was transferred and resulted in a singleton pregnancy with delivery.
D.4 Ghost PNs (absent NPBs)/single NPB in one or more PNs (bull’s eye)
Despite the difficulty of making comparative studies aimed at evaluating the relevance of PN scoring, the sequence of processes involved in fertilization underlines the importance and significance of NPBs and nucleoli in determining embryo viability. In light of these considerations, it is not surprising that the absence of nucleoli in PNs at the time of fertilization check, the so-called ‘ghost’ PNs (Figs 181–184), or the presence of a single NPB, known as ‘bull's eye’ PNs (Figs 185–189) has been reported to be associated with epigenetic defects and abnormal development in animals (Svarcova et al., 2009).
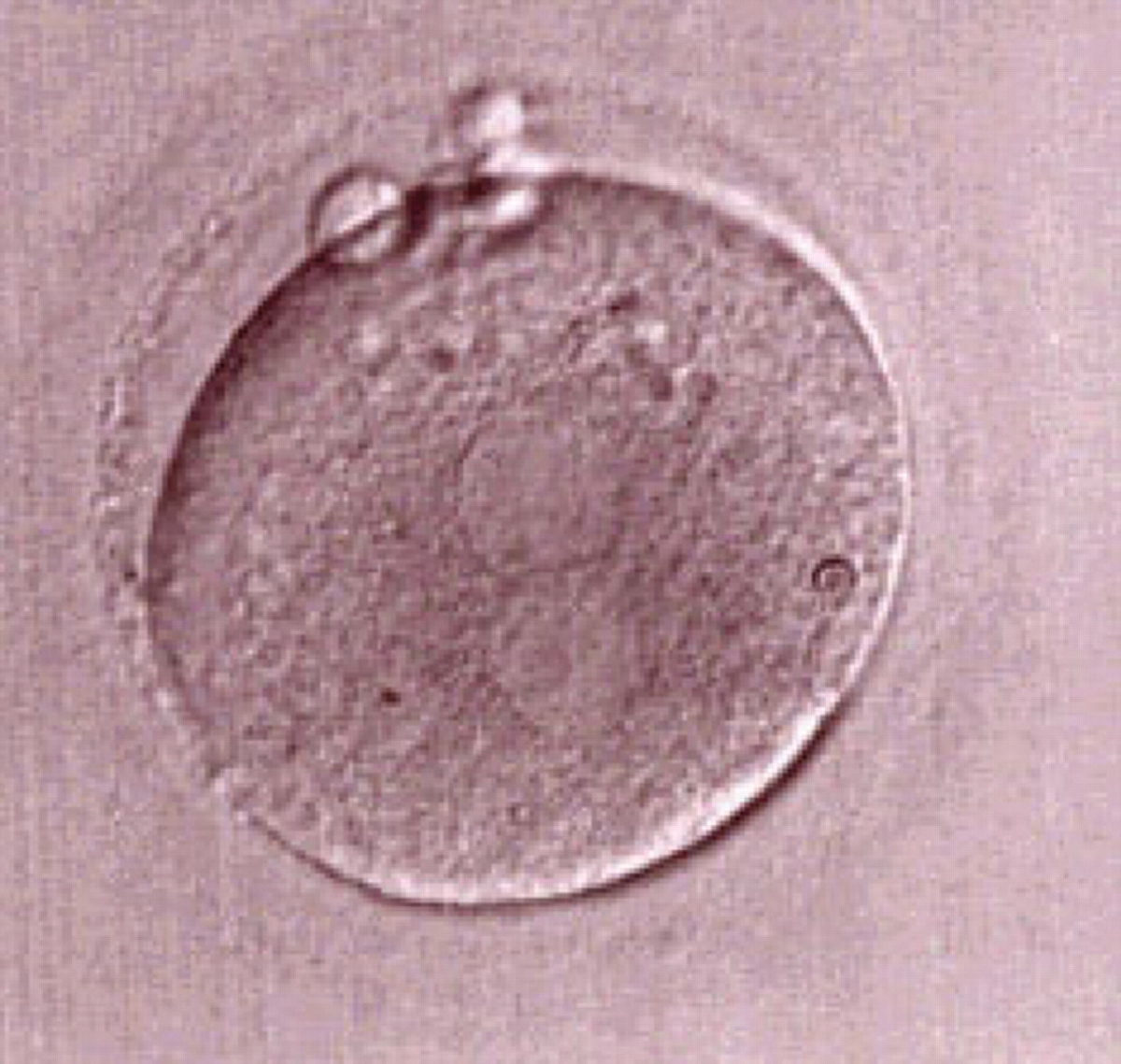
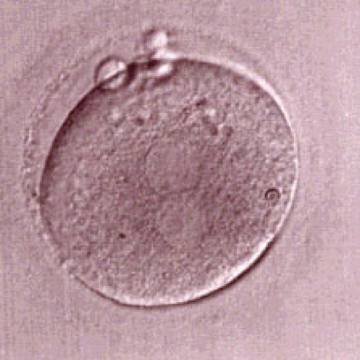
Figure 181
One out of three sibling zygotes (see also Figs 182 and 183) generated by ICSI using frozen/thawed epididymal sperm observed 18 h post-insemination. All three zygotes clearly show two distinct PNs with well-defined membranes and without any NPBs. All three were transferred and two healthy baby boys were
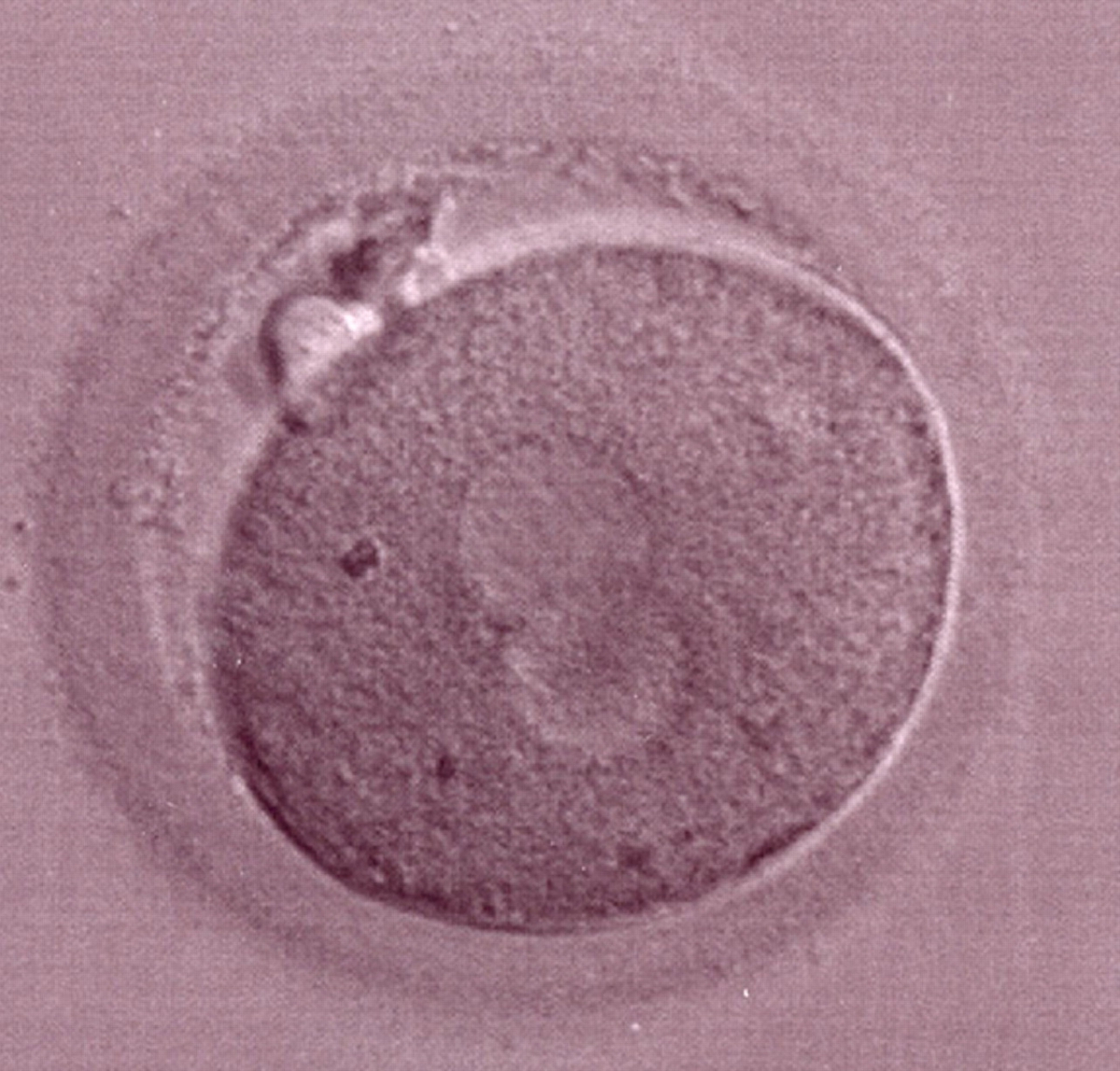
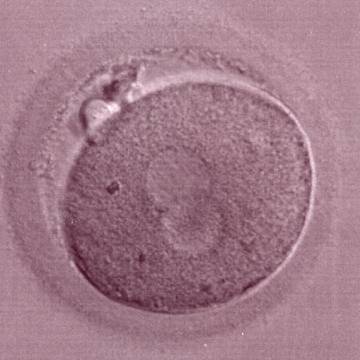
Figure 182
The second out of three sibling zygotes (see also Figs 181 and 183) generated by ICSI using frozen/thawed epididymal sperm. All three zygotes showed refractile bodies in the cytoplasm.
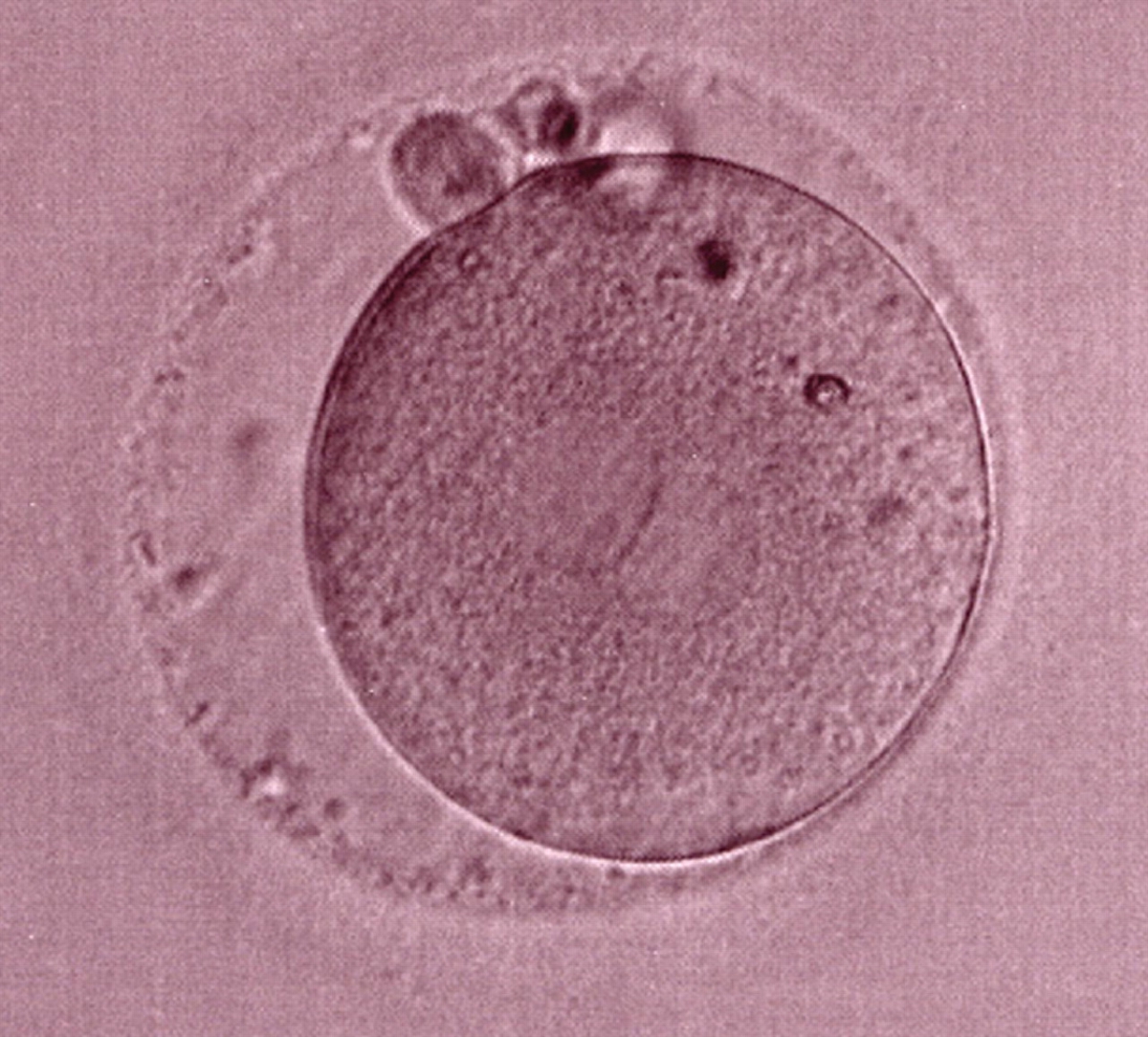
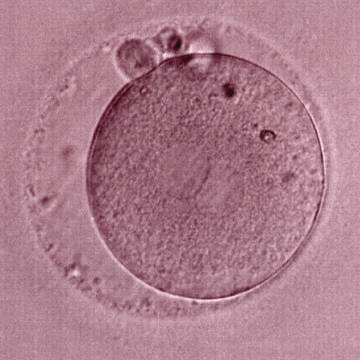
Figure 183
The third of three sibling zygotes (see also Figs 181 and 182) generated by ICSI using frozen/thawed epididymal sperm. Beside the absence of NPBs and the presence of refractile bodies, this zygote has a large perivitelline space.
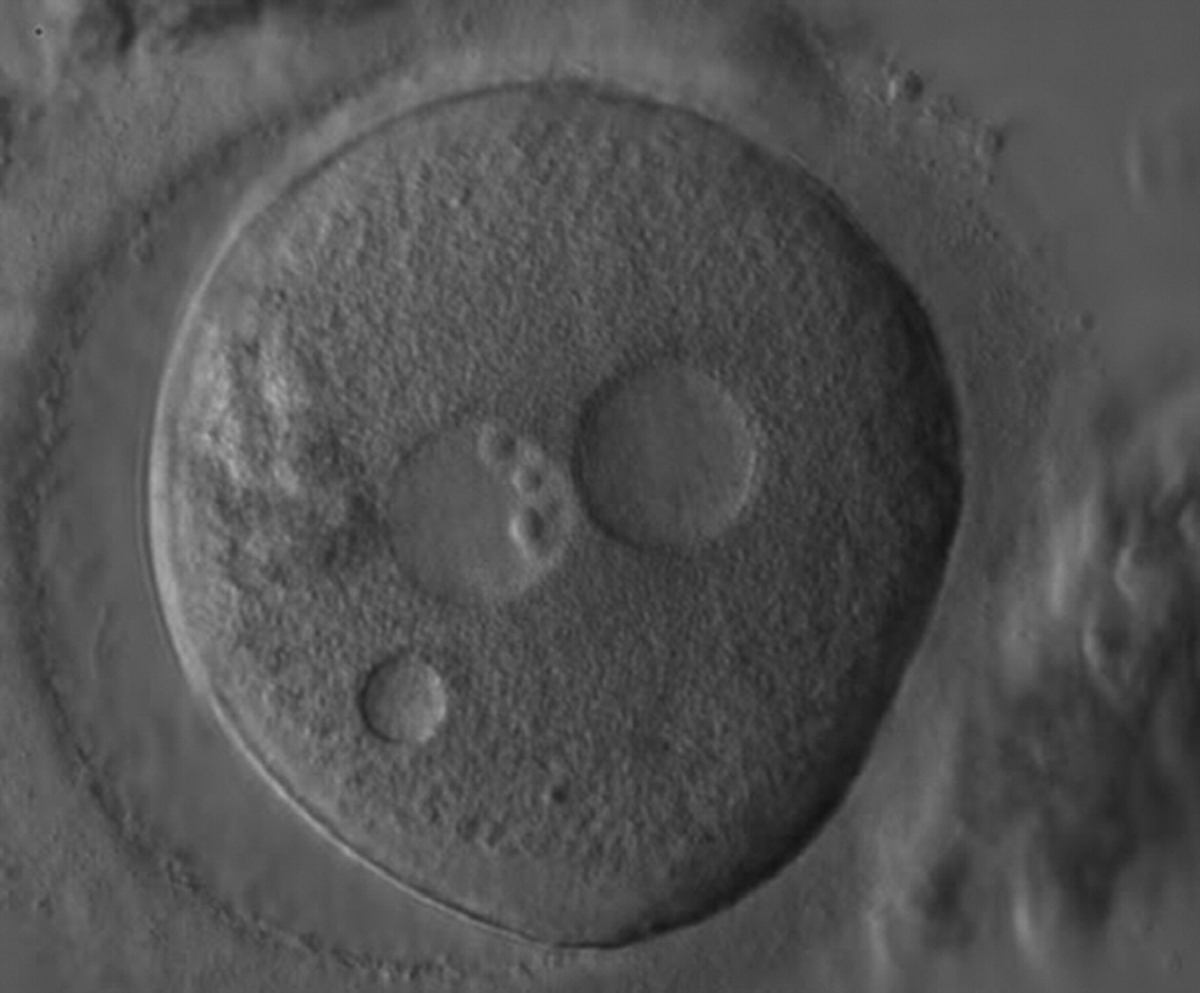
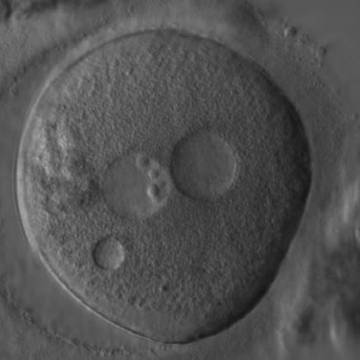
Figure 184
A zygote generated by ICSI showing what looks like two distinct PNs with distinct membranes and the absence of NPBs in one of the PNs (400× magnification). One small vacuole is visible under the left PN. Two highly fragmented polar bodies are present at 9 o'clock. Despite the presence of 2 polar bodies after ICSI, it is possible that this is a 1PN zygote, and that the structure to the right is a vacuole (compare with Fig. 209). It was not transferred.
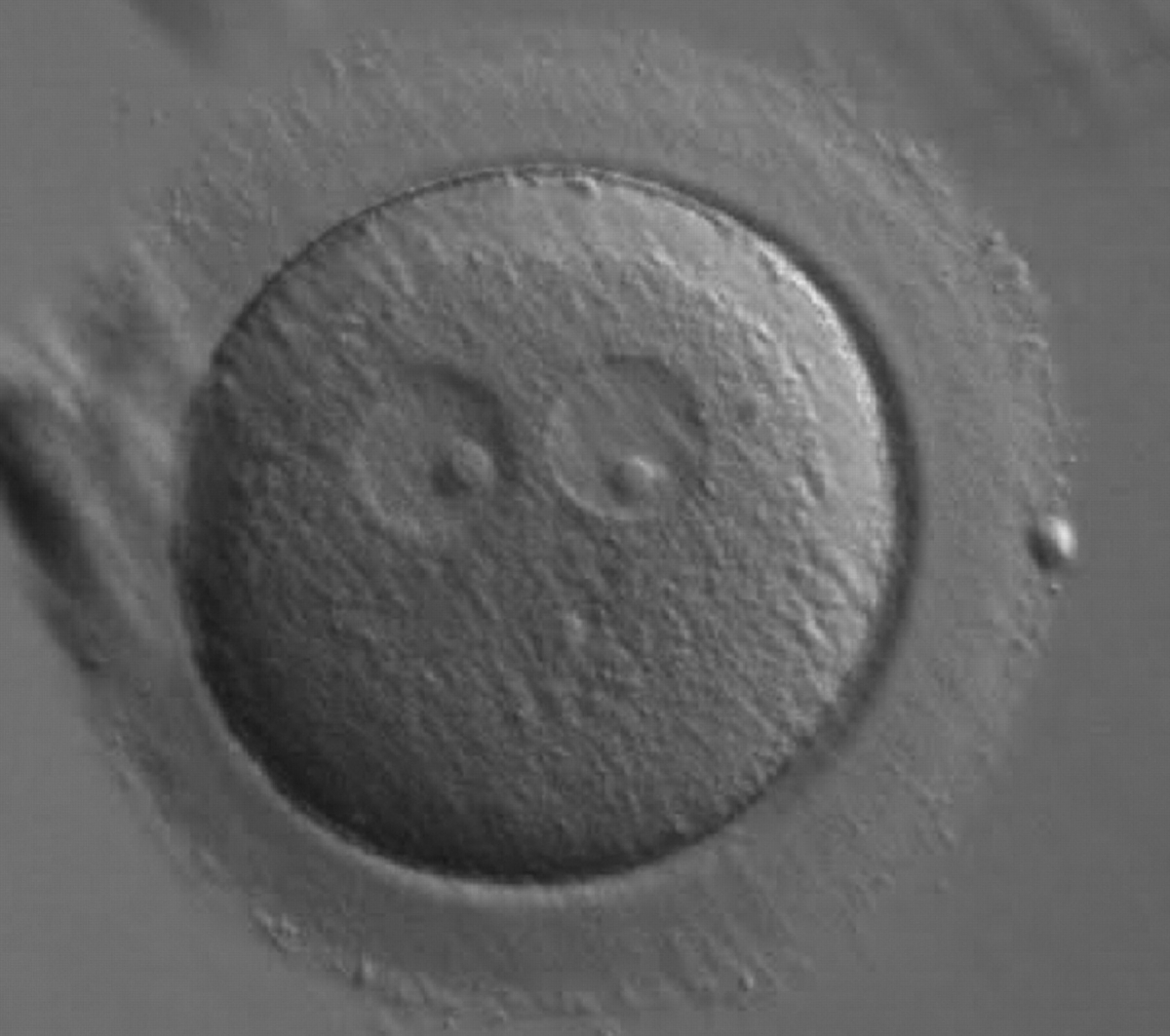
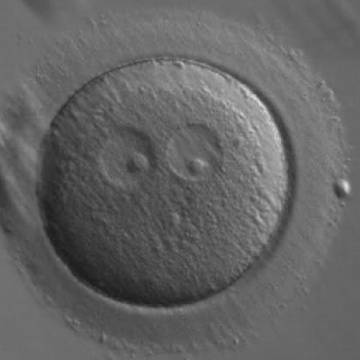
Figure 185
A zygote generated by ICSI showing two ‘bull's eye’ PNs, each having a single large NPB (200× magnification). The PNs are slightly separated and are not as yet juxtaposed. A clear cortical region is evident in the cytoplasm. It was discarded.
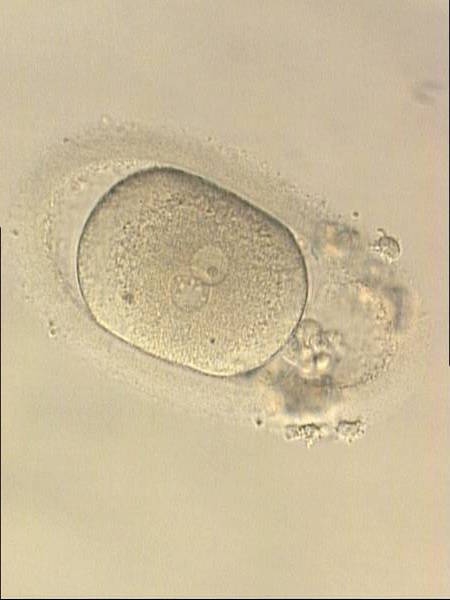
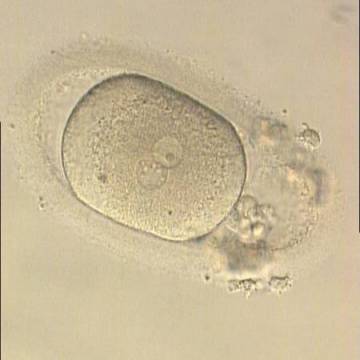
Figure 186
Oval-shaped zygote generated by ICSI, showing a single NPB in one of the two PNs (‘bull's eye’) and small, scattered NPBs in the other (200× magnification). The PVS is quite large and the polar bodies are highly fragmented. It was discarded.
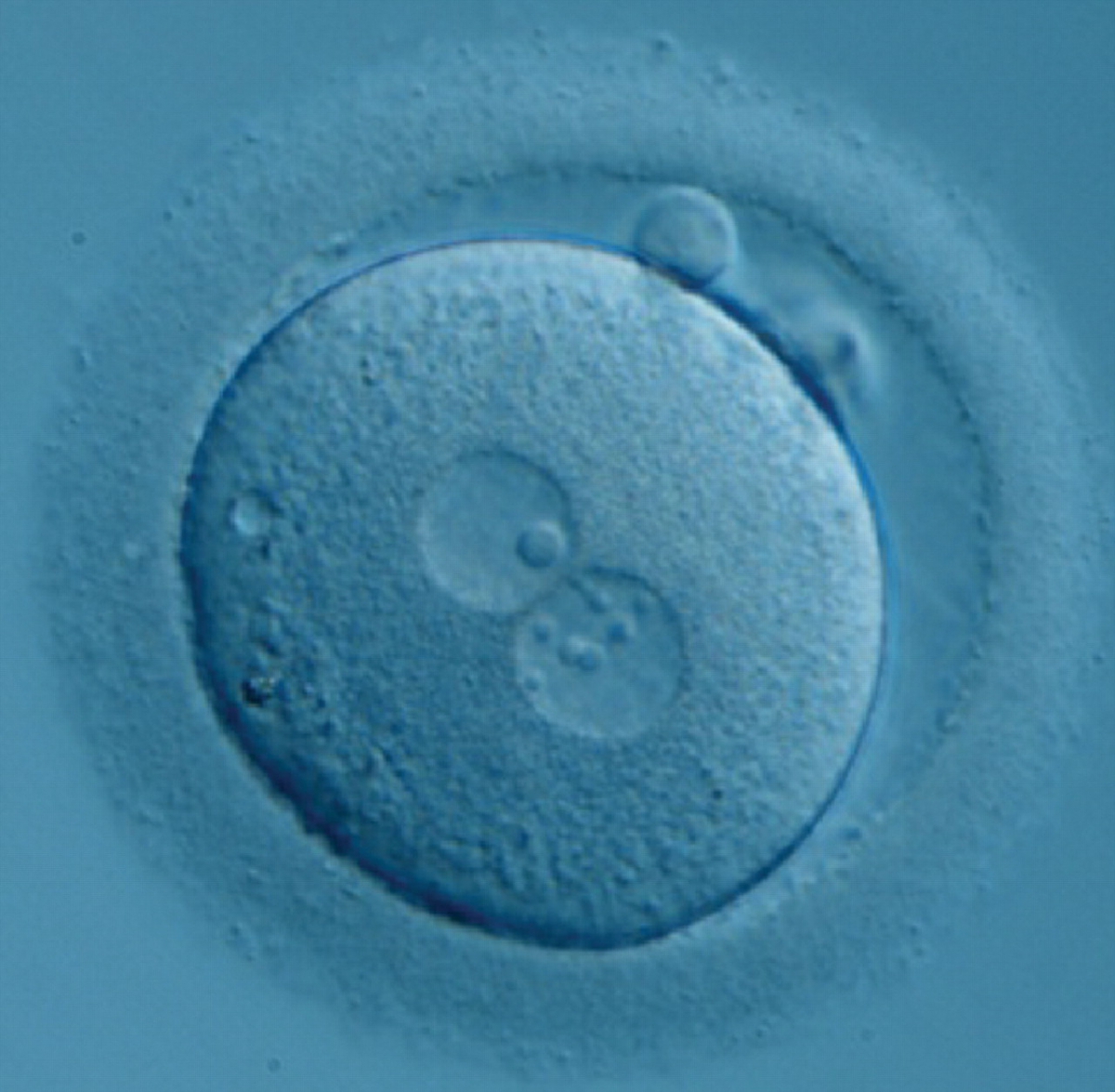
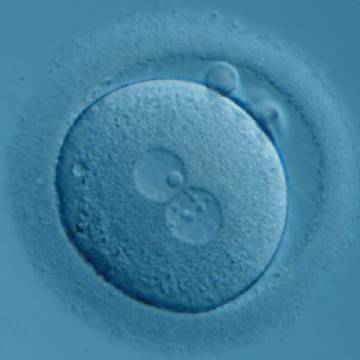
Figure 187
Zygote generated by ICSI with one ‘bull's eye’ NPB in one PN and small, different-sized NPBs in the other (400× magnification). It was transferred but clinical outcome is unknown.
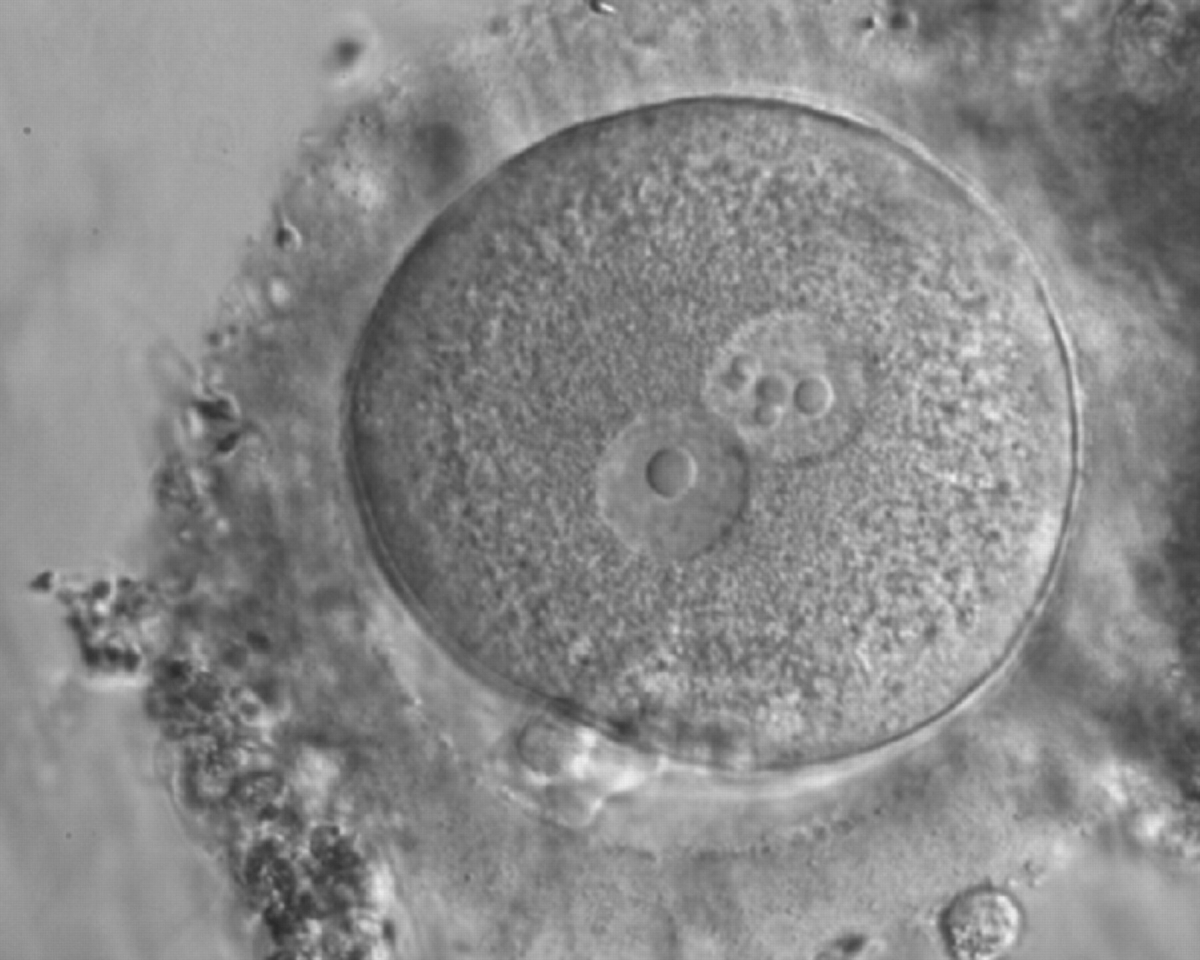
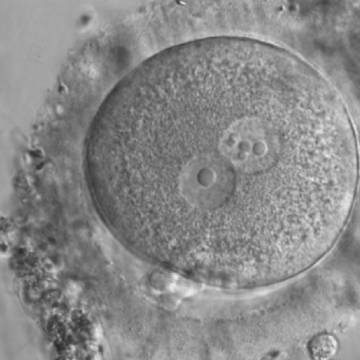
Figure 188
Zygote generated by ICSI displaying two centrally positioned PNs: in one PN there is a single large NPB (‘bull's eye’), whereas the NPBs are smaller, different-sized and scattered in the other (600× magnification). It was discarded.
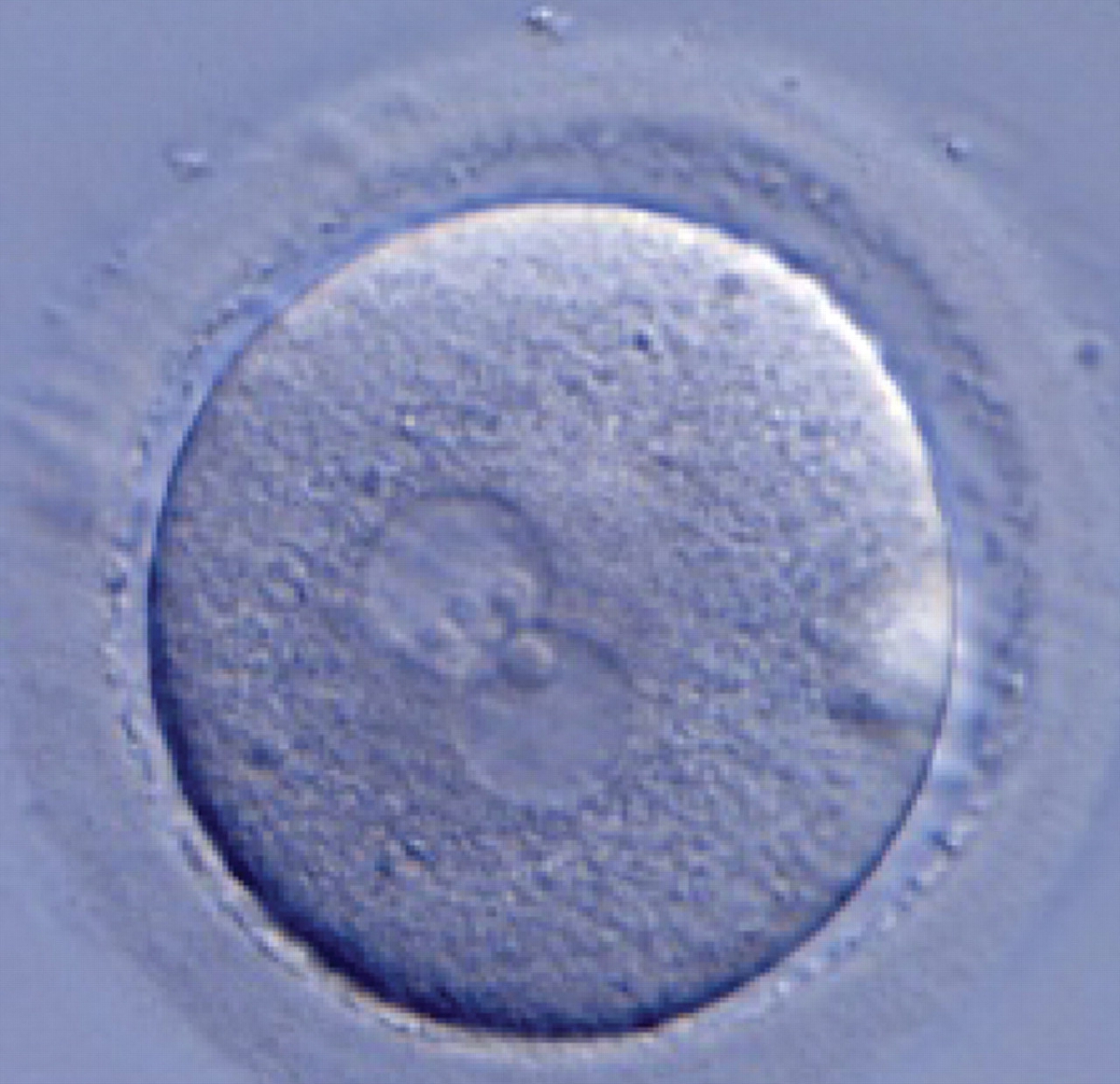
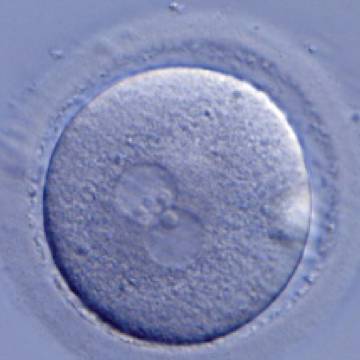
Figure 189
A zygote generated by ICSI with one ‘bull's eye’ PN (400× magnification). The NPBs from each PN are aligned at the PN junction. It was transferred but clinical outcome is unknown.
Article references:
Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum Reprod 2011;26:1270-1283.
Abstract/FREE Full Text
Balaban B, Urman B, Isiklar A, Alatas C, Aksoy S, Mercan R, Mumcu A, Nuhoglu A. The effect of pronuclear morphology on embryo quality parameters and blastocyst transfer outcome. Hum Reprod 2001;16:2357-2361.
Abstract/FREE Full Text
Brezinova J, Oborna I, Svobodova M, Fingerova H. Evaluation of day one embryo quality and IVF outcome—a comparison of two scoring systems. Reprod Biol Endocrinol 2009;7:9.
CrossRef | Medline | Google Scholar
Ebner T, Moser M, Sommergruber M, Gaiswinkler U, Wiesinger R, Puchner M, Tews G. Presence, but not type or degree of extension, of a cytoplasmic halo has a significant influence on preimplantation development and implantation behaviour. Hum Reprod 2003;18:2406-2412.
Abstract/FREE Full Text
Edwards RG, Beard HK. Oocyte polarity and cell determination in early mammalian embryos. Mol Hum Reprod 1997;3:863-905.
Abstract/FREE Full Text
James AN, Hennessy S, Reggio B, Wiemer K, Larsen F, Cohen J. The limited importance of pronuclear scoring of human zygotes. Hum Reprod 2006;21:1599-1604.
Abstract/FREE Full Text
Montag M, van der Ven H. Evaluation of pronuclear morphology as the only selection criterion for further embryo culture and transfer: results of a prospective multicentre study. Hum Reprod 2001;16:2384-2389.
Abstract/FREE Full Text
Montag M, Liebenthron J, Köster M. Which morphological scoring system is relevant in human embryo development? Placenta 2011;32:S252-S256.
CrossRef | Medline | Web of Science | Google Scholar
Nicoli A, Valli B, Di Girolamo R, Di Tommaso B, Gallinelli A, La Sala GB. Limited importance of pre-embryo pronuclear morphology (zygote score) in assisted reproduction outcome in the absence of embryo cryopreservation. Fertil Steril 2007;88:1167-1173.
CrossRef | Medline | Web of Science | Google Scholar
Payne D, Flaherty SP, Barry MF, Matthews CD. Preliminary observations on polar body extrusion and pronuclear formation in human oocytes using time-lapse video cinematography. Hum Reprod 1997;12:532-541.
Abstract/FREE Full Text
Payne JF, Raburn DJ, Couchman GM, Price TM, Jamison MG, Walmer DK. Relationship between pre-embryo pronuclear morphology (zygote score) and standard day 2 or 3 embryo morphology with regard to assisted reproductive technique outcomes. Fertil Steril 2005;84:900-909.
CrossRef | Medline | Web of Science | Google Scholar
Pedersen T. Growth factors in the nucleolus? J Cell Biol 1998;143:279-281.
FREE Full Text
Scott LA, Smith S. The successful use of pronuclear embryo transfers the day following oocyte retrieval. Hum Reprod 1998;13:1003-1013.
Abstract/FREE Full Text
Scott L, Alvero R, Leondires M, Miller B. The morphology of human pronuclear embryos is positively related to blastocyst development and implantation. Hum Reprod 2000;15:2394-2403.
Abstract/FREE Full Text
Scott L. Oocyte and embryo polarity. Semin Reprod Med 2001;18:171-183.
Google Scholar
Scott L. Pronuclear scoring as a predictor of embryo development. Reprod Biomed Online 2003;6:201-214.
CrossRef | Medline | Google Scholar
Senn A, Urner F, Chanson A, Primi MP, Wirthner DW, Germond M. Morphological scoring of human pronuclear zygotes for prediction of pregnancy outcome. Hum Reprod 2006;21:234-239.
Abstract/FREE Full Text
Svarcova O, Dinnyes A, Polgar Z, Bodo S, Adorjan M, Meng Q, Maddox-Hyttel P. Nucleolar re-activation is delayed in mouse embryos cloned from two different cell lines. Mol Reprod Dev 2009;76:132-141.
CrossRef | Medline | Web of Science | Google Scholar
Tesarik J, Greco E. The probability of abnormal preimplantation development can be predicted by a single static observation on pronuclear stage morphology. Hum Reprod 1999;14:1318-1323.
Abstract/FREE Full Text
Van Blerkom J, Antczak M, Schrader R. The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum Reprod 1997;12:1047-1055.
Abstract/FREE Full Text
